Keywords
Cryodamage; Sperm; Boar; Mitochondrial activation; Mitochondrial superoxide; ROS production; Lipid peroxidation
Introduction
For artificial insemination (AI), boar semen is prepared in rather simple extenders, kept at 16-20°C and usually used in liquid form for up to 3 days from extension. Routine freezing of boar semen is occasionally used for gene banking, replacement of lost genetics or long-distance transport [1]. The reason behind this meagre use of freezing is the low tolerance of the ejaculated boar sperm to current methods used which, consequently, leads to lower conception rates and smaller litter sizes after AI, compared with fresh semen [2]. While freezing leads to severe physical changes in osmolality due to asymmetric ice formation and deleterious concentration of solutes [3] thus affecting cell survival, simple cooling cause’s oxidative stress mainly produced for the generation of reactive oxygen species (ROS) [4]. In this regard, lipoperoxidation of membrane lipids could lead to membrane destabilization, directly affecting sperm survival post-thaw but also the function of the surviving sperm. The high sensitivity to cooling (cold-shock) depicted by boar sperm probably owes to their peculiar lipid composition of the sperm membrane, rich in palmitic acid (C16:0), stearic acid (C18:0) and oleic acid (C18:1 n-9) as well as about 25% docosapentaenoic acid (C22:5 n-6; DPA) and 30% docosaheaenoic acid (C22:6 n-3; DHA) [5,6]. Ejaculated sperm lose membrane cholesterol at various events such as capacitation [7] and dramatically during cryopreservation [8], which affects the surviving sperm. Sperm survival has been related to the ratios of DPA and DHA to the other membrane fatty acids, owing to the beneficial effects of fewer long-chain polyunsaturated fatty acids (PUFA) against an increase in saturated fatty acids (SFA) [8], provided an eternal source of phospholipids is available. Less is known about the role of lecithin glycerophospholipids phosphatydil-choline (LPC) or phosphatydiletanolamine (LPE), which compose 20% respectively 8-20% of all lecithin’s, or even the less abundant phosphatydil-serine (LPS). LPC and LPE build drops associated with their membrane while LPS seems to attach to washed boar sperm [9], but whether they relate to avoidance of oxidative stress on sperm membrane remains to be tested. Soybean lecithin, with almost 100% of LPC, has been successfully used for freezing sperm as substitute of egg yolk in several species at different concentrations: 3% (human [10]), 1-3% (dog [11]), 1.5% (rabbit [12]), 3.5% (ram [13]), 2-5% (25% LPC, effective concentration: 0.5-1.25%; brown bear [14]) but boar, exception made of an study using LPC (0.2%) to evaluate the effect of semen refrigeration [15].
Boar epidydymal sperm seems more resistant to freezing that ejaculated sperm, for reasons yet unknown [16]. Aquaporin 7, a water channel membrane protein that is differentially located in cauda and ejaculated sperm might be related to this phenomenon [17]. Boar sperm also appear to acquire cryo-resistance if kept in seminal plasma (SP) at room temperature for up to 4 h postejaculation [9], probably due to the attachment of seminal plasma exosomes and cholesterol lipoproteins that are able to diminish exit of cholesterol from the sperm membrane [18]. However, while the SP is usually removed, a historical addition of the undefined egg-yolk (20% v/v) has proven protective enough against cooling damage and, when used in combination with glycerol, to further protect from freezing damage [19] by replacing the destabilizing loss of cholesterol from the sperm membrane. Egg yolk is thus not only commonly used for the cryopreservation of the semen of boar [20] but also for other domestic (ram [13]; bull [21]) and wild species (brown bear [22]). Use of egg yolk faces, however, problems due to its varying composition, its undefined chemistry and its animal origin.
Since the egg yolk used in most extenders binds to the sperm plasma membrane and can replace cholesterol and membrane phospholipids [23], specific components have been studied in many species but porcine: (a) lipoproteins (high and low density) associated to cholesterol and (b) lecithin (glycerophospholipids). The low-density lipoprotein (LDL) is postulated as the main responsible for the cryoprotectant activity and it has been used as a suitable replacement of whole egg yolk in different studies (reviewed by Wang et al., [24]). Lyophilized lipoprotein fractions isolated from egg yolk have been previously tested [25,26]. The assumption is taken that the effect of an increased adsorption of phospholipids to the membrane is enhanced by the emulsifying action of extension detergents, rich in lauryl sulfate (as OEP or Orvus WA), which are customarily included in egg yolk extenders. However, too high concentrations or lengthy exposures of/to lauryl sulfate are able to disrupt the sperm plasma membrane [27].
In the present study, we tested the hypothesis that chemically defined phospholipids could replace egg yolk in the cooling extender during freezing of sperm exposed (ejaculated) or not (cauda epididymis) to seminal plasma, from the same boars, ameliorating the deleterious effects of conventional cryopreservation, with a particular focus on oxidative stress. Two different types of phospholipids were tested: (i) natural: phosphatidyl-choline from egg yolk (LPCE) and from soybean (LPCS) lecithin and (ii) synthetic: phosphatidyl-glycerol (LPGE). To the best of our knowledge, this is the first study using specific phospholipids to freeze boar sperm.
Material and Methods
Reagents
All reagents were obtained from Sigma-Aldrich (Sweden), unless otherwise stated. Phospholipids (Lipoids), phosphatidyl choline from egg lecithin (CAS-No. 97281-44-2), phosphatidyl choline from soybean lecithin (CAS-No. 97281-47-5), phosphatidyl glycerol from egg lecithin (CAS-No. 92347-24-5) were kindly provided by Lipoid AG, Switzerland (www.lipoid.com). Fluorochomes for flow cytometry were pursued from Molecular Probes, INVITROGEN: YO-PRO-1 Iodide (YO-PRO-1), PNA-AleaFluor488 (Arachis hypogaea Lectin (Peanut)-AleaFluor488), Propidium Iodide (PI,), MitoTracker Deep Red (MIT), MitoSOX Red (MSO), CellROX Green (CellROX), SYTO Red Dead Cell Stain (SYTO), 4,4-difluoro- 5-(4-phenyl-1,3-butadienyl)-4-bora-3a,4a-diaza-s-indacene- 3-undecanoic acid - BODIPY 581/591 C11 (BODIPY-C11), JC-1 (JC-1) except Hoechst 33342 (H33342, used in all the stainingcombinations to define DNA-containing events and discard the debris), Sigma-Aldrich, Sweden.
Animals, sperm collection
Mature Swedish Landrace breeding boars (n=5) selected according to normal semen quality and proven fertility were used. Boars were individually kept on straw beds at the Centre for Biomedical Resources (CBR, LiU). All animals were fed with commercial rations (Läntmännen, Stockholm, Sweden) according to national standards [28] and provided with water ad libitum.
Ejaculated sperm were obtained by manual (gloved-hand method) collection of the sperm-rich fraction (SRF) of the ejaculate, twice weekly. Only ejaculates with at least 70% motile and 75% morphologically normal sperm immediately after collection were used. The SRF-sperm were extended in Beltsville-Thawing Solution (BTS, IMV-Technologies, L´Aigle, France) (1:4 extension), cooled to 16-20°C and transported to the laboratory for freezing.
Epididymis were collected from the same boars (n=5) after euthanasia by exsanguination under general anesthesia. Epididymal sperm were obtained by serially cutting the caudal epididymis duct with a scalpel, collecting the fluid into sterile 10 mL plastic tubes. The epididymal sperm were thereafter washed twice in BTS to avoid eventual blood contamination, and extended in BTS (1:4 dilutions), cooled to 16-20°C and transported to the laboratory for freezing.
All experiments were performed at the Department of Clinical and Experimental Medicine, Linköping University, Linköping, Sweden in accordance with relevant regulations (European Community Directive 2010/63/EU) and compliance with Swedish current legislation (SJVFS 2012:26). The Experimental protocol had previously been reviewed and approved by the Local Ethical Committee for Experimentation with Animals, at Linköping (permit nr 74-12), Sweden.
Sperm freezing
The BTS-extended sperm were centrifuged (300 × g/ 10 min, room temperature) and the resulting pellet split into four aliquots. Each aliquot was re-suspended with each refrigeration extender (Control: LEY; Experimental: LPCE, LPCS and LPGE, see refrigeration extender), to a final concentration of 1.5 × 109 sperm/ml. All samples were slowly cooled (≈ -0.25°C/min to +5°C) before being mixed with a single freezing extender (see freezing extender) containing 9% glycerol (Control samples also containing 1.5% Orvus WA (963-1000, Preservation Equipment Ltd, UK)), at a ratio of two parts of semen to one part of extender to reach a final concentration of 1 × 109 sperm/mL and 3% glycerol rate (Control samples 0.5% OWA final concentration). Sperm were then packaged at +5°C 0.5 mL plastic straws, and frozen in a programmable bio-freezer (IceCube 14 M-A; Minitube International, Tiefenbach, Germany) at -6°C/min from +5°C to -5°C, holding for crystallization 1 min at -5°C, at -60°C/min from -5°C to -140°C. The straws were then plunged into liquid nitrogen (LN2, -196°C) for storage until thawing for analyses, minimum 1 week. The straws were thawed in circulating water at 38°C for 15 seconds and diluted directly 1:4 in BTS pre-warmed (38°C).
Refrigeration and freezing extender: Ejaculated sperm:
I Control. LEY: 80% β -lactose 310 mM (L310) + 20 % egg yolk.
II LPCE: L310 + LIPOID E PC (0.5 % - LPCE1; 1 % - LPCE2; 1.5 % - LPCE3; 2 % - LPCE4; and 4 % - LPCE5 - w/v). Phosphatidyl choline from egg lecithin (≥ 98.0 g Phosphatidyl choline/100 g product). CAS-No. 97281-44-2.
III LPCS: Lactose 310 mM + LIPOID S PC (0.5 % - LPCS1; 1 % - LPCS2; 1.5 % - LPCS3; 2 % - LPCS4; and 4 % - LPCS5 - w/v). Phosphatidyl choline from soybean lecithin (≥ 98.0 g Phosphatidyl choline/100 g product). CAS-No. 97281-47-5.
IV LPGE: Lactose 310 mM + LIPOIDS E PG (0.5 % - LPGE1; 1 % - LPGE2 and 2 % - LPGE3 - w/v). Phosphatidyl glycerol from egg lecithin (≥ 98.0 g Phosphatidyl glycerol/100 g product). CAS-No. 92347-24-5.
Epididymis sperm: Only Control, LPCE 2, LPCS2 and LPGE1 were used for cryopreservation of cauda epidydimal sperm, according to post-thawing results from ejaculated sperm.
Freezing extenders were prepared adding 9 % glycerol (I to IV) plus 1.5 % OWA (only Control), for final concentrations of 3% glycerol and 0.5% OWA.
Sperm assessment
Sperm motility: Sperm total motility, progressive motility and velocity were assessed using an upright Zeiss Aio Scope A1 light microscope equipped with a 10 phase contrast objective (Carl Zeiss, Stockholm, Sweden) connected via a CMOS camera (UEye, IDS Imaging Development Systems GmbH, Ubersulm, Germany) to a computer holding the QualispermTM software (Biophos SA, Lausanne, Switzerland). Semen droplets (24 × 104 sperm in10 μL) were placed on a pre-warmed Menzel-Gläser pre-cleaned microscope slide (ThermoFisher Scientific, Waltham, MA, USA. Size 76 26 mm) covered by a pre-warmed coverslip (Size 18 18 mm, VWR, Stockholm, Sweden), on a thermal plate (Temp Controller 2000-2, Pecon GmbH, Erbach, Germany) kept at 38 ºC. The QualispermTM technology is based on fluorescence correlation spectroscopy analysis of single particles (sperm) in confocal volume elements, yielding a regression fluctuation algorithm. Individual sperm are projected on a pixel grid of the CMOS camera and the algorithm calculates the number of fluctuations in each pixel by correlation function of sperm numbers and translation classes. From the correlation function the speed (velocity) distribution is then determined. This system benefits from a high throughput (usually 4 fields per minute), analyzing >2,000 sperm/field and has been validated for several species, including porcine [29].
Flow cytometry: Stock solutions of the fluorochromes were prepared in miliQ water (PI and H33342), and in Dimethyl sulfoxide (DMSO, SIGMA-ALDRICH, Sweden) (YO-PRO-1, MIT, MSO, BODIPY-C11, JC-1). CellROX/SYTO kit contains ready to use solutions. All Stock solutions were kept at -20°C (except at +4°C for H33342) and brought to RT immediately before diluting them in BTS for its use. Working solutions of the fluorochromes were prepared in BTS: 4.5 μM H33342, 1 μg/mL PNA-AleaFluor488, 2.4 μM PI, 200 nM YO-PRO-1, 100 nM MIT, 5 μ M MSO, 4.5 μM BODIPYC11, 1.5 nM JC-1, 500 nM CellROX and 5 nM SYTO. Sperm suspensions (1 × 106 sperm/mL) were analyzed in a Gallios™ flow cytometer (Beckman Coulter, Bromma, Sweden) equipped with standards optics, violet laser (405 nm) 2 colours, argon laser (488 nm) 5 colours and HeNe-laser (633 nm) 3 colours. Filter configuration: Blue: FL1 550SP 525BP (YO-PRO-1, PNAAleaFluor488, CellROX, BODIPYC11), FL2 595SP 575BP (JC-1, MSO, BODIPYC11), FL3 655SP 620/30 (PI, SYTO), FL4 730SP 695/30 - alt 675BP, FL5 755LP; Red: FL6 710SP 660BP (MIT), FL7 750SP 725/20, FL8 755LP; Violet: FL9 480SP 450/50 (H33342), FL10 550/40. The instrument is controlled via Navios software (Beckman Coulter, Bromma, Sweden). Analyses of acquired data were performed using the Kaluza software (Beckman Coulter, Bromma, Sweden) on a separate PC. In all cases we assessed 25,000 events per sample, with a flow rate of 500 cells/sec.
I Viability and acrosome status (H33342/PI/PNA-AleaFuor488): Sperm viability and acrosome status were assessed using the triple stain PNA-AleaFluor488, PI and H33342, yielding the percentage of sperm viable and with intact acrosomes (Viab; PI-PNA-AleaFluor488-).
II Membrane integrity, early membrane destabilization changes, mitochondrial superoxide indicator and mitochondrial status (H33342/YO-PRO-1/PI/MT/MSO): Membrane integrity and early membrane destabilization changes were assessed using YO-PRO-1 and PI, yielding the percentage of viable sperm with the membrane intact (Membrane destabilization; PI-YO-PRO-) and of sperm viable and with early membrane destabilization (Membrane destabilization; PI- YO-PRO+). Mitotracker Deep Red (MIT) was used to address the mitochondrial status. The flow cytometry analysis yielded, from the percentage of sperm viable, sperm with active mitochondria (Mitochondrial activation; PI- MIT+) and inactive mitochondria (PI-MIT-). For the mitochondrial superoxide status, the MitoSOX mitochondrial superoxide indicator was used. The analysis yields, from the percentage of sperm viable, those sperm positive for superoxide-oxidation (Mitochondrial superoxide production; PI- MSO+).
III Membrane mitochondrial potential (H33342/JC-1): Membrane mitochondrial potential was assessed using JC-1. This probe accumulates in the mitochondria green-fluorescence monomer. In the presence of high mitochondrial membrane potential (Δψm), the monomers form aggregates, shifting the fluorescence to orange. The FC analysis typically shows three populations, from events with high orange and low green fluorescence (JC- 1; JC-1 orange: high Δψm;) to events with low orange and high green fluorescence (low Δψm) and an intermediate population of medium orange and high green fluorescence (medium Δψm).
IV Oxidative stress detection (H33342/CellROX/SYTO): Generation of reactive oxygen species under conditions of oxidative stress, including cryopreservation process, could be detected by using CellROX in combination with SYTO Red Dead Cell. The analysis yields, from the percentage of live sperm, those positive for ROS production (ROS production; CellROX+ SYTO-).
V Lipid peroxidation (H33342/BODIPY-C11): The probe BODIPY-C11 was used to assess the degree of LPO-positive cells (Lipid peroxidation; BODIPY-C11+). The probe emits orange fluorescence in its non-iodized state, shifting to green when peroxidized.
Statistical analysis
The variables collected were statistically analyzed in the R statistical environment (R Development Core Team, R Foundation for Statistical Computing, Vienna, Austria. ISBN 3-900051-07- 0). Results are shown as means and standard error of the mean (SEM), unless otherwise stated. Normal distribution of the data was analyzed using the Shapiro-Wilk test and further analyses of the post-thawing data were carried out using linear mixed-effects models, with transformation of data (arcsin) in case of nonnormal distribution (total and progressive motility). Two models were built with type of extender as fied effects, and (1) ejaculated or (2) cauda epididymal sperm as the random part of the model (with or without exposure to SP). Significant fixed effects were further analyzed using multiple comparisons of means with Tukey contrasts. An additional model (inter-source analysis) was built in order to compare the 4 specific extenders used in both sperm sources (ejaculates and cauda epididymal sperm collected from the same animals).
Results
Osmotic pressure
The osmotic pressure measured in the frozen samples (sperm+extender) post-thaw did not vary among groups (Table 1).
| |
Cooling |
PT-EJ |
PT-EP |
| Extender |
| LEY |
367 |
804±34 |
846 ± 46 |
| LPCE0.5 |
348 |
694±49 |
- |
| LPCE1 |
341 |
704 ± 19 |
- |
| LPCE1.5 |
343 |
740±10 |
- |
| LPCE2 |
360 |
758±3 |
810 ± 55 |
| LPCE4 |
361 |
874 ± 23 |
- |
| LPCS0.5 |
345 |
712 ± 37 |
- |
| LPCS1 |
345 |
785 ± 13 |
- |
| LPCS1.5 |
345 |
799 ± 6 |
- |
| LPCS2 |
368 |
817 ± 23 |
846 ± 55 |
| LPCS4 |
445 |
825 ± 22 |
- |
| LPGE0.5 |
345 |
690 ± 23 |
- |
| LPGE1 |
371 |
748 ± 10 |
810 ± 32 |
| LPGE2 |
449 |
757 ± 16 |
- |
Table 1: Osmotic pressure in the sperm extenders used (mOsm/kg), extender without glycerol (Cooling Extender), post-thawed ejaculated sperm (PT-EJ) and post-thawed epididymis sperm (PT-EJ) containing 3% glycerol.
Sperm motility post-thaw
Post-thawed total and progressive motility of ejaculated sperm (EJ) in all treatments were significantly lower than control (Table 2). Although sperm velocity followed the same pattern among treatments, the LPCS2 remained noteworthy similar to control. Moreover, control in ejaculated were lower that epididymis (intersource analysis). All motility variables post-thaw for epididymal sperm (EP) were significantly lower (p<0.05) than controls (Table 2).
| Ejaculated |
TM |
PM |
Vel |
| LEYG 0 |
34 ± 7.7 a |
31.5 ± 7.7 a |
17.6 ± 1.6 a |
| LPCE 0.5 |
1.5 ± 1.5* |
1.2 ± 1.2* |
4.5 ± 4.5* |
| LPCE 1 |
0 ± 0* |
0 ± 0* |
0 ± 0* |
| LPCE 1.5 |
1.3 ± 1.3* |
0.9 ± 0.9* |
4.5 ± 4.5* |
| LPCE 2 |
2.3 ± 0.9* b |
1.6 ± 0.5* b |
5.6 ± 0.5* b |
| LPCE 4 |
0 ± 0* |
0 ± 0* |
0 ± 0* |
| LPCS 0.5 |
1 ± 1* |
1 ± 1* |
2.7 ± 2.7* |
| LPCS 1 |
0 ± 0* |
0 ± 0* |
0 ± 0* |
| LPCS 1.5 |
0 ± 0* |
0 ± 0* |
0 ± 0* |
| LPCS 2 |
9.5 ± 3* b |
8.5 ± 3.2* b |
17.4 ± 4.7 ab |
| LPCS 4 |
0 ± 0* |
0 ± 0* |
0 ± 0* |
| LPGE 0.5 |
0 ± 0* |
0 ± 0* |
0 ± 0* |
| LPGE 1 |
2.8 ± 1* b |
2.3 ± 0.8* b |
5.8 ± 1.7* b |
| LPGE 2 |
0 ± 0* |
0 ± 0* |
0 ± 0* |
| Epididymis |
| LEYG 0 |
47.8 ± 7.4 a |
45.2 ± 7.6 a |
29.7 ± 2.8 c |
| LPCE 2 |
8.5 ± 3.7* b |
7.5 ± 3.5* b |
19.2 ± 2.7* ac |
| LPCS 2 |
6.3 ± 2.7* b |
4.6 ± 2.1* b |
11.1 ± 3.2* b |
| LPGE 1 |
0.2 ± 0.2* b |
0.2 ± 0.2* b |
1.5 ± 1.5* b |
Table 2: Post-thaw motility (%) and sperm velocity (microm/sec) of cauda and ejaculated boar spermatozoa frozen with emulsified crude egg-yolk (control) or various specific phospholipids in the extender, Total motility (TM), progressive motility (PM) and velocity (Vel) were analyzed. Results are shown as means and standard errors of the mean (Mean ± SEM),*Indicates differences (p<0.05), relative to the control -LEYGO, for each parameter, for each origin of the sample (ejaculated or epididymis). Letters indicates differences (p<0.05) among extenders for both sample sources.
Sperm membrane, mitochondrial and oxidative status
Sperm viability: Membrane integrity (Viab) in EJ (p<0.05) showed lower values for all groups relative to the control, except for LPCE2, LPCS2 and LPGE1, whereas no differences were found in EP (Table 3). The cauda epididymal sperm control group (emulsified egg yolk) depicted lower sperm viability than ejaculated sperm control group (p<0.05).
Early membrane destabilization changes: The percentage of sperm with an intact membrane and that of sperm alive with early membrane destabilization changes showed no differences, relative with control for neither EJ nor EP.
Mitochondrial activation, superoxide production and membrane potential in mitochondria: Proportions of high membrane potential (Mit potential; JC-1) were lower for all the groups in EJ (p<0.05), relative to the control (Table 3) and only significant for LPGE1 in EP. The inter-source analysis showed no differences between controls, whereas LPCE2 and LPCS2 values in ejaculated samples were lower than for cauda epididymal sperm (p<0.05). In addition, mitochondrial activation in EJ was lower in all the groups (except LPCE2 and LPCS2) (p<0.05), results that are repeated in EP samples (Figure 1) and also in the inter-source analysis (Supplementary Table 1). Regarding the mitochondrial superoxide production in the mitochondrion (MitoSOX) in EJ (Figure 1), higher percentages (p<0.05) of oxidative stress (PI−/ MISO+) were found in LPCE1.5, LPCS0.5 and LPGE1 and LPGE2, relative to the control (p>0.05) (Supplementary Table 1). This parameter in EP was only significant for the LPGE1 (Figure 1), but not in the inter-source analysis (Supplementary Table 1).
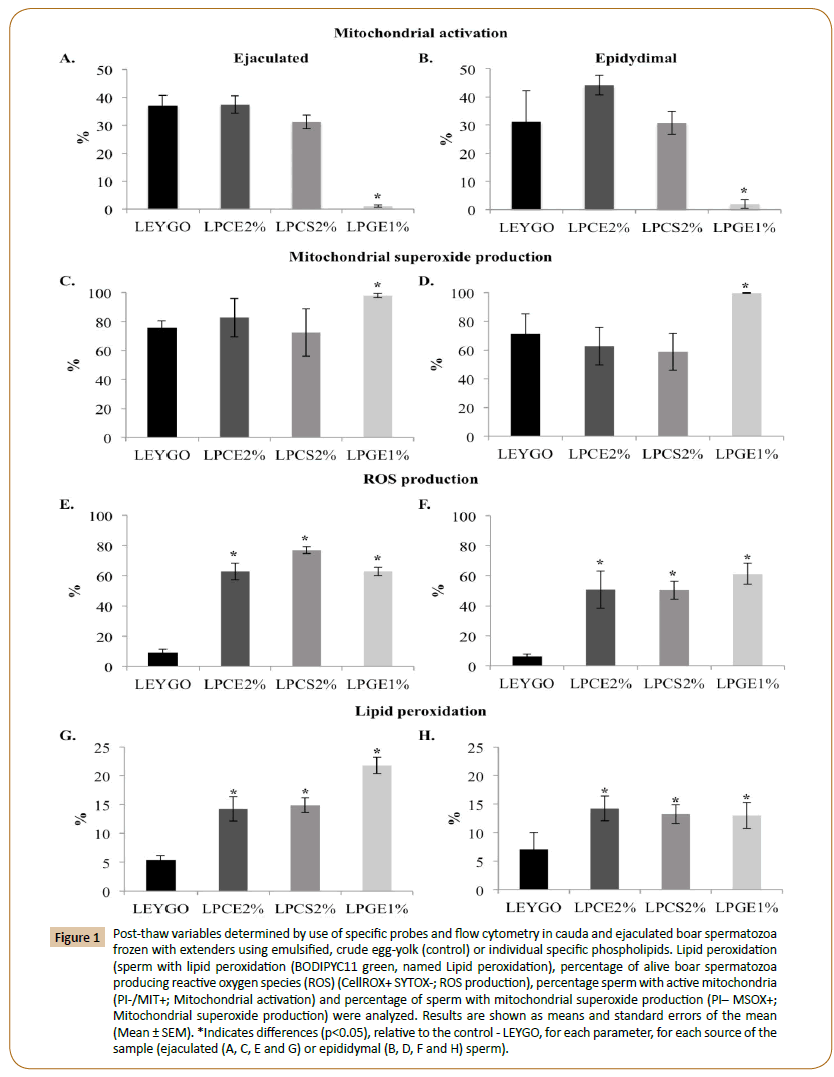
Figure 1: Post-thaw variables determined by use of specific probes and flow cytometry in cauda and ejaculated boar spermatozoa frozen with extenders using emulsified, crude egg-yolk (control) or individual specific phospholipids. Lipid peroxidation (sperm with lipid peroxidation (BODIPYC11 green, named Lipid peroxidation), percentage of alive boar spermatozoa producing reactive oxygen species (ROS) (CellROX+ SYTOX-; ROS production), percentage sperm with active mitochondria (PI-/MIT+; Mitochondrial activation) and percentage of sperm with mitochondrial superoxide production (PI– MSOX+;Mitochondrial superoxide production) were analyzed. Results are shown as means and standard errors of the mean (Mean ± SEM). *Indicates differences (p<0.05), relative to the control - LEYGO, for each parameter, for each source of the sample (ejaculated (A, C, E and G) or epididymal (B, D, F and H) sperm).
| Ejaculated |
Viab |
Membraneintegrity |
Membranedestabilization |
Mitpotential |
| LEYGO |
52.6 ± 3.6 a |
45.2 ± 2.9 |
17.9 ± 7.7 |
33.7 ± 3.4 a |
| LPCE0.5 |
21.6 ± 7.9* |
15.9 ± 1.0* |
4.7 ± 1.5 |
24.8 ± 9.9* |
| LPCE1 |
23.2 ± 4.7* |
13.6 ± 4.8* |
26 ± 0 |
14.5 ± 4.3* |
| LPCE1.5 |
18 ± 7.8* |
15.1 ± 3.0* |
2.7 ± 1.3 |
19.8 ± 8* |
| LPCE2 |
42.1 ± 4.2 b |
38.9 ± 3.7* |
6.2 ± 0.9 |
13.4 ± 1.6* b |
| LPCE4 |
13.5 ± 8.8* |
24.9 ± 9.0* |
16.3 ± 0 |
10.8 ± 4.5* |
| LPCS0.5 |
21 ± 4.6* |
18.1 ± 5.7* |
4.2 ± 1 |
19.6 ± 4* |
| LPCS1 |
24.9 ± 6.6* |
17 ± 6.5* |
21.5 ± 0 |
15.3 ± 5.4* |
| LPCS1.5 |
17.9 ± 5.4* |
11.2 ± 3.5* |
5.2 ± 0 |
11.7 ± 1.6* |
| LPCS2 |
43.4 ± 1.3 b |
34.6 ± 3.6* |
7.9 ± 3 |
11.2 ± 1.4* b |
| LPCS4 |
7.8 ± 3.5* |
19 ± 6.0* |
23.5 ± 0 |
7.3 ± 5* |
| LPGE0.5 |
34 ± 8.4* |
23.8 ± 5.8* |
22.6 ± 0 |
9.7 ± 5.4* |
| LPGE1 |
47.6 ± 5.2 ab |
50.1 ± 3.1* |
8.2 ± 1 |
10.6 ± 1.6* b |
| LPGE2 |
16.3 ± 9.5* |
9.3 ± 6.0* |
13.4 ± 0 |
5.3 ± 0.8* |
| Epididymal |
| LEYGO |
43.1 ± 5.2 b |
37.3 ± 9.3 |
22.7 ± 7.4 |
35.6 ± 4.9 ac |
| LPCE2 |
48.9 ± 2.8 ab |
46.2 ± 2.9 |
10.3 ± 3.7 |
33.5 ± 0.5 ac |
| LPCS2 |
41.1 ± 2.4 b |
32.1 ± 3.8 |
18.9 ± 2.7 |
23.3 ± 1.5 cd |
| LPGE1 |
44.7 ± 4.9 ab |
35.7 ± 5.7 |
7.7 ± 0.2 |
19.7 ± 6.5* bd |
Table 3: Flow cytometry variables assessed post-thaw in cauda and ejaculated boar spermatozoa frozen with emulsified crude egg-yolk (control) or individual specific phospholipids in the extender, Viab: Sperm viability (PI-), Membrane integrity: sperm viability with intact membrane (YO-PRO-1-/PI-), Membrane destabilization: early membrane destabilization changes (YO-PRO-1+/PI-), Mit potential: membrane mitochondrial potential (JC-1; JC-1 orange). Results are shown as means and standard errors of the mean (Mean±S.E.M). *Indicates differences (p<0.05), relative to the control – LEYGO, for each parameter, for each origin of the sample (ejaculated or epididymal sperm). Letters indicates differences (p<0.05) among extenders for both sample sources.
Lipid peroxidation: Lipid peroxidation, monitored with the BODIPYC11-probe, was higher in all treatment groups relative to controls, both in EJ and EP (p<0.05) (Figure 1G and 1H). Controls did not show significant differences (Supplementary Table 1).
Oxidative stress: Percentages of alive sperm positive for oxidative stress (ROS production) (Figure 1E and 1F), was higher in both EJ and EP compared to controls, results confirmed in the intersource analysis (Supplementary Table 1).
Correlation analysis of sperm kinematics and flow cytometry parameters: In EJ, high positive correlations were found between kinetic variables and mitochondrial potential, mitochondrial activation and membrane integrity. Interestingly, there was a negative correlation between TM and PM with lipid peroxidation and ROS production (Table 4). Thus, an increased TM for an individual boar might negatively mirror oxidative stress among surviving sperm (lipid peroxidation and ROS production). In EP, the correlations were fewer, but yet displaying a similar pattern: sperm motility of surviving sperm correlates with the levels oxidative stress caused by cooling. Overall, the R-values were higher in the ejaculated sperm for membrane integrity, mitochondrial potential and mitochondrial activation. In contrast, the correlation between ROS production and motility parameters was higher for cauda epididymal sperm; whereas it was lower for the rest of the studied parameters. A similar correlation pattern was observed for lipid peroxidation.
| |
PM |
TM |
Vel |
Membraneintegrity |
Viab |
Membranedestabilization |
Mitpotential |
MitochondrialActivation |
Mitochondrialsuperoxideproduction |
Lipidperoxidation |
ROS production |
|
| PM |
|
0.9988*** |
0.6575*** |
0.5809*** |
0.6426*** |
-0.074 |
0.6324*** |
0.7096*** |
-0.0682 |
-0.5709*** |
-0.5736*** |
EJACULATED |
| TM |
0.9988*** |
|
0.6799*** |
0.5961*** |
0.6500*** |
-0.0668 |
0.6436*** |
0.7253*** |
-0.0863 |
-0.5868*** |
-0.5913*** |
| Velocity |
0.8161*** |
0.8232*** |
|
0.6621*** |
0.6538*** |
-0.0771 |
0.6305*** |
0.7179*** |
-0.0568 |
-0.6453*** |
-0.5085*** |
| Membraneintegrity |
-0.0389 |
-0.0619 |
0.1224 |
|
0.8417*** |
-0.2507 |
0.5482*** |
0.7304*** |
0.0873 |
-0.3833** |
-0.3996** |
| Viab |
-0.1294 |
-0.1437 |
0.02 |
0.491 |
|
-0.1798 |
0.6494*** |
0.6776*** |
-0.0049 |
-0.5189*** |
-0.3858** |
| Membranedestabilization |
0.5263* |
0.5389 |
0.1658 |
-0.4695 |
-0.3346 |
|
-0.1705 |
-0.1594 |
-0.2966* |
-0.1493 |
-0.3130* |
| Mitpotential |
0.4007 |
0.4028 |
0.5970* |
0.6069* |
0.4871 |
0.0366 |
|
0.6051*** |
-0.1209 |
-0.5930*** |
-0.5192*** |
| MitochondrialActivation |
0.1919 |
0.1921 |
0.5609* |
0.6124* |
0.4103 |
-0.1175 |
0.6115* |
|
-0.1334 |
-0.5574*** |
0.4656*** |
| Mitochondrialsuperoxideproduction |
-0.0877 |
-0.1085 |
-0.2905 |
0.1663 |
0.3933 |
-0.0596 |
-0.0768 |
-0.2732 |
|
0.2839* |
0.1458 |
| Lipidperoxidation |
-0.5592* |
-0.5432* |
-0.3163 |
-0.3503 |
-0.0394 |
-0.3629 |
-0.3706 |
-0.2285 |
-0.4397 |
|
0.6746*** |
| ROS production |
-0.7556*** |
-0.7546*** |
-0.6745** |
-0.1368 |
-0.036 |
-0.406 |
-0.494 |
-0.3027 |
0.1238 |
0.4483 |
|
| |
EPIDIDYMAL |
|
Table 4: Correlations (Pearson) (r-values) between variables determined post-thaw in ejaculated or cauda epididymal spermatozoa retrieved from the same boars, Total motility (TM), progressive motility (PM), velocity (Vel), sperm viability with intact membrane (YO-PRO-1-/PI-; Membrane integrity), sperm viability (PI-; Viab), early membrane destabilization changes (YO-PRO-1+/PI-; Membranedestabilization), membrane mitochondrial potential (JC-1 orange; Mit potential), sperm with active mitochondrion (PI-/MIT+; Mitochondrial activation) and superoxide production in the mitochondria (PI– MSOX+; Mitochondrial superoxide production), sperm with lipid peroxidation (BODIPYC11 green; Lipid peroxidation), production of reactive oxygen species (ROS) by alive boar spermatozoa (CellROX+ SYTOX–; ROS production) were analysed. Results are shown as r-values and “-”indicates negative correlations. *p<0.05; **p<0.01; ***p<0.001.
Discussion
Despite documented efforts to reach acceptable fertility and prolificacy after AI in pigs, the overall cryosurvival of ejaculated semen is still consistently lower than in other species, most probably due to the cellular damage occurring during conventional cryopreservation [30]. Empirical addition of different cryoprotectants and antioxidants has been tested, with variable effects, yet in all cases the cryo-survival being low [1]. Comparison of ejaculated and cauda epididymal sperm has shown that motility, viability and membrane mitochondrial potential were superior for post-thawed cauda epididymal sperm; perhaps due to their lower intracellular H2O2 production [16], related to the differential location of Aquaporin 7 between sources [17]. Sperm ffreezing and thawing induces a decrease in membrane fluidity with considerable inter-male variation [31], probably caused by the different amounts of long-chain PUFA in the sperm plasma membrane, among sires [32]. Oxidative attack to this PUFA, mainly in terms of lipid peroxidation, could be corrected by physiological levels of ROS, thus aiding preservation of normal sperm function [33]. In our study, mitochondrial superoxide production reached very high levels for all tested groups, compared to controls, where both ROS production and lipid peroxidation were reduced. Whether specific changes on sperm membrane composition and sperm quality after cryopreservation are related to other specific components of the egg yolk than phospholipids remains to be elucidated. This scenario, added to the use of ill-defined hen egg yolk, has defined the current methods as sub-optimal. To this sub-optimality we need to add that the methods used for analysis of the cryopreserved sperm have mostly focused on cryo-survival indicators (i.e., motility, membrane integrity) and only in few cases of experimental nature were the surviving sperm screened in more detail, both for intactness of their chromatin, their mitochondrial function and the reactivity of their plasma lemma, over time. These overtime studies are increasingly relevant in terms of sperm functionality after insemination [18].
In the present study, two different types of individual phospholipids were tested as replacement of the customary OWA-emulsified egg yolk in both ejaculated sperm and also cauda epididymal sperm from the very same boars: i) natural: phosphatidyl-choline from egg yolk and from soybean lecithin (0.5 to 4% concentration) and ii) synthetic: phosphatidyl-glycerol (0.5 to 2%); comparing the individual sole use of these specific phospholipids against a control etender using OEP-emulsified eggyolk at 20%. Glycerol was used as cryoprotectent in either case. The choice of these range of concentrations for the individual phospholipids were defined by the composition of egg yolk: 20% of egg-yolk contained approximately 1.35% of phosphatidylcholine [34] and the soybean extenders: from soybean lecithin, 19-21% is phosphatidyl-choline [35].
The replacement of crude egg yolk by low-density lipoproteins (LDL) resulted in higher cryosurvival after thawing [25]. Use of low-density lipoproteins (LDLs), most often isolated from eggyolk from different species has proven beneficial for sperm function post-thaw, particularly to maintain DNA-integrity [36]. In contrast, our present results clearly showed that solely using specific phospholipids in an individual layout was not able to maintain the sperm quality shown by the control extender (egg yolk-based), measured by different analytical techniques postthaw. In a recent study, Wang et al. [24] concluded that 0.09 g/mL (9%) of LDL could replace egg yolk for freezing boar sperm. In this regard, the LDL fraction, which accounts for approximately twothirds of the material in egg yolk, has been ascribed responsible for the effect [37]. In LDL, only 28% are phospholipids, being 73% of these PC [33]. Therefore, the equivalence of PC contained in 9% LDL accounts to approx. 0.5%. However, single use of such a concentration in the present study was not able to maintain similar values as when using crude egg yolk, irrespective of the parameter considered. The specific LDL-component phosphatidylcholine has been successfully used at 1% concentration (10 mg/ mL) to improve the function of turkey semen stored at 4°C for 24 h in terms of lipid peroxidation levels [38]. In contrast, our results showed that none of the concentrations tested (0.5 to 4%) could maintain the levels neither of lipid peroxidation nor of ROS production, both for ejaculated and cauda epididymal sperm at controls levels. Even if these differences might relate to the species tested, there is a possibility that a single phospholipid, irrespective of concentration is not sufficient for a full protection of membrane intactness in pigs. It might be so that other specific components of the lecithin, contained in the LDL, are key actors in the stabilization of the membrane, as shown by the use of 1.5% soybean lecithin (ram [39]).
The cryopreservation process severely impairs mitochondrial activity in boar [40], as we could confirm in our study that only about one third of the mitochondria remain actives postthawing (control group). Specific phospholipids can elicit paradoical effects. For instance, PG is a surfactant agent that, in combination with Ca2+, may induce perturbations in the bilayer structures by formation of structures aggregates [41]. Being PG a constituent molecule of the mitochondrial membrane, the inclusion of this phospholipid in the cryopreservation extender can induce a significant decrease in mitochondrial activation and an increase in mitochondrial superoxide production; both events being registered in the present study, alongside decreases in sperm motility post-thaw. Specific interactions with the sperm membrane maybe affect the membrane fluidity, most-likely modifying the cholesterol balances. Fraser et al. [42] concluded that lipoprotein fraction, presumably phospholipids, confers protection to the sperm membrane during boar liquid storage. In our study, individual use of phospholipids did not prevent the detrimental effect of cryopreservation so that maybe a combined action is required for such a complex preservation mechanism. Therefore, reconsideration is needed when deciding which phospholipid is to be added, and to which concentration.
Mitochondria maintained activity post-thaw better among control and phosphatidyl-choline treated ejaculated sperm than in cauda epididymal sperm. Independent of the extender used, freezing/thawing lead to superoxide formation in the mitochondria, reaching in the control extender, levels of up to 75%. This is not surprising, since accompanying effects on the fluidity of membranes, including that of the acrosome and the mitochondria, were clearly seen when lipid peroxidation was assessed in freshly stored and frozen/thawed sperm using mass spectrometric analysis of the main endogenous lipid classes, phosphatidyl-choline and cholesterol and by fluorescence techniques using the lipid peroxidation reporter probe C11- BODIPY (581/591) [43]. It is known that lipids from cryopreserved cells contained significantly less sphingomyelin (14.6 +/- 1.1 vs 22.4 +/- 1.6%) and more phosphatidyl-choline (51.5 +/- 2.0 vs 40.5 +/- 2.4%) [44]. The high susceptibility to cryoprotectants such as glycerol depicted by boar sperm could be linked to its particular membrane composition, which may affect cryosurvival; as suggested by the high levels of superoxide production seen in our study, even in control samples. In contrast, emulsified eggyolk was able to maintain levels of ROS production and lipid peroxidation low, probably linked to the existence of specific protective compounds in the whole egg-yolk, whose existence is yet to be proven. Replacement by isolated phospholipids at the rates tested in the present study does not seem enough to avoid cryodamage.
Exposure of sperm to seminal plasma (SP) before extension and cooling has been considered beneficial for cryopreservation [45], while other authors found that its influence is detrimental before freezing [46]. Post-thaw eposure to SP yield beneficial effects; increase of motility and reduction of chromatin alterations [47], increasing sperm survival [48] and even increased conception rates [45]. Several studies have defined the higher “freezability” of cauda epididymal samples [16] and even the higher results of in vitro fertilization obtained post-thawing [49], in comparison with ejaculated samples. However, our inter-source comparison of samples derived from the very same boars showed the cauda epididymal sperm were less viable than the ejaculated ones. We can only speculate on the reasons behind these evident differences.
In the present study, both cauda epidydimal fluid and SP were presumably removed by double washing of the sperm suspensions with BTS. Washing has, however, not proven the most effective method to remove these protein-rich fluids that are quickly adsorbed to the sperm plasma membrane [50] so that the interaction with the added phospholipids could be impaired, or the membrane be less affected by the non-inclusion of the laurylsulphate, compared to controls. Moreover, the lack of differences among the rest of parameters studied (except sperm velocity, which was highest in epidydimal sperm) could suggest that the removal of the protective cauda epididymal fluid be contra productive, in relation to the ejaculated SP. Interestingly, the motion-related parameter mitochondrial membrane potential, was also higher in cauda epididymal sperm, for both PC sources. In addition, the relation of these two parameters (velocity and membrane potential) was highly positively correlated (p<0.05). This differential effect ought to be more deeply revised.
Conclusion
Our results showed that the addition of individual phospholipids, isolated from egg yolk or soybean lecithin, were under the present Experimental layout using a conventional freezing method, unable to prevent cryodamage of neither ejaculated nor did epididymal (cauda) boar sperm, compared to an extender using 20% emulsify crude egg-yolk. Further studies about the combination of different phospholipids and also the combined effect with lecithin are needed to assay their eventual protective effect, particularly in relation to the source of sperm.
Conflict of Interest
The authors declare no conflict of interest.
Author Contribution
MAR performed the experiments and wrote the first draft of the manuscript. AVC assisted with the experiments and draft writing. HRM designed the experiments, supervised the work and corrected the manuscript. All authors approved the final version of the manuscript.
Acknowledgements
Authors acknowledge Lipoids.com Company for kindly provided the different phospholipids used. The study has been made possible by grants from The Swedish Research council VR, Stockholm (Grant 521-2011-6553), the Research Council FORMAS (Grant 221-2011-512), Stockholm, and FORSS (Forskningsrådet i Sydöstra Sverige, Grant 473121), Sweden.
References
- Yeste M (2016)Sperm cryopreservation update: Cryodamage, markers, and factors affecting the sperm freezability in pigs. Theriogenology 85: 47-64.
- Roca J, Parrilla I, Bolarin A, Martinez EA, Rodriguez-Martinez H (2016) Will AI in pigs become more efficient? Theriogenology 86: 187-93.
- Mazur P (1984) Freezing of living cells: mechanisms and implications. Am J Physiol 247: 125-142.
- Aitken RJ (1989) The role of free oxygen radicals and sperm function. Int J Androl12: 95-97.
- Penny PC, Noble RC, Maldjian A, Cerolini S (2000) Potential role of lipids for the enhancement of boar fertility and fecundity. Pig News and Information 25: 119-126.
- Rooke JA, Shao CC, Speake BK (2001) Effects of feeding tuna oil on the lipid composition of pig spermatozoa and in vitro characteristics of semen. Reproduction 121: 315-322.
- Flesch FM, Brouwers JF, Nievelstein PF, Verkleij AJ, van Golde LM, et al. (2001) Bicarbonate stimulated phospholipid scrambling induces cholesterol redistribution and enables cholesterol depletion in the sperm plasma membrane. J Cell Sci 114: 3543-3555.
- Maldjian A, Pizzi F, Gliozzi T, Cerolini S, Penny P, et al. (2005) Changes in sperm quality and lipid composition during cryopreservation of boar semen. Theriogenology 63: 411-421.
- Butler WJ, Roberts TK (1975) Effects of some phosphatidyl compounds on boar spermatozoa following cold shock or slow cooling. J Reprod Fertil 43: 183-187.
- Vireque AA, Tata A, Silva OF, LoTurco EG, Azzolini A, et al. (2016) Effects of n-6 and n-3 polyunsaturated acid-rich soybean phosphatidylcholine on membrane lipid profile and cryotolerance of human sperm. Fertil Steril 106: 273-283.
- Axnér E, Lagerson E (2016) Cryopreservation of Dog Semen in a Tris Extender with 1% or 2% Soya Bean Lecithin as a Replacement of Egg Yolk. Reprod Domest Anim 51: 262-268.
- Nishijima K, Kitajima S, Koshimoto C, Morimoto M, Watanabe T, et al. (2015) Motility and fertility of rabbit sperm cryopreserved using soybean lecithin as an alternative to egg yolk. Theriogenology 84: 1172-1175.
- Mata-Campuzano M, Alvarez-Rodríguez M, Alvarez M, Tamayo-Canul J, Anel L, et al. (2015) Post-thawing quality and incubation resilience of cryopreserved ram spermatozoa are affected by antioxidant supplementation and choice of extender. Theriogenology 83:520-528.
- Alvarez-Rodríguez M, Alvarez M, Anel-López L, Martínez-Rodríguez C, Martínez-Pastor F, et al. (2013) The antioxidant effects of soybean lecithin- or low-density lipoprotein-based extenders for the cryopreservation of brown-bear (Ursus arctos) spermatozoa. Reprod Fertil Dev 25: 1185-1193.
- Simpson AM, Swan MA, White IG (1987) Susceptibility of epididymal boar sperm to cold shock and protective action of phosphatidylcholine. Gamete Res 17: 355-73.
- Alkmin DV, Perez-Patiño C, Barranco I, Parrilla I, Vazquez JM, et al. (2014) Boar sperm cryosurvival is better after exposure to seminal plasma from selected fractions than to those from entire ejaculate. Cryobiology 69: 203-210.
- Vicente-Carrillo A, Ekwall H, Alvarez-Rodriguez M, Rodriguez-Martinez H (2016) Membrane Stress During Thawing Elicits Redistribution of Aquaporin 7 But Not of Aquaporin 9 in Boar Spermatozoa. Reprod DomestAnim 51: 665-679.
- https://www.intechopen.com/articles/show/title/cryopreservation-of-pig-spermatozoa-oocytes-and-embryos-state-of-the-art
- Lovelock JE, Polge C (1954) The immobilization of spermatozoa by freezing and thawing and the protective action of glycerol. Biochem J 58: 618-622.
- Hu JH, Li QW, Jiang ZL, Li WY (2008) Effects of different extenders on DNA integrity of boar spermatozoa following freezing–thawing. Cryobiology 57: 257-262.
- Amirat L, Tainturier D, Jeanneau L, Thorin C, Gérard O, et al. (2004) Bull semen in vitro fertility after cryopreservation using egg-yolk LDL: a comparison with Optidyl, a commercial egg-yolk extender. Theriogenology 61: 895-907.
- Anel L, Gomes-Alves S, Alvarez M, Borragan S, Anel E, et al. (2010) Effect of basic factors of extender composition on post-thawing quality of brown bear electroejaculated spermatozoa. Theriogenology 74: 643-651.
- Cerolini S, Maldjian A, Pizzi F, Gliozzi TM (2001) Changes in sperm quality and lipid composition during cryopreservation of boar semen. Reproduction 121: 395-401.
- Wang P, Wang YF, Wang CW, Bu SH, Hu JH, et al. (2014) Effects of low-density lipoproteins extracted from different avian yolks on boar spermatozoa quality following freezing-thawing.Zygote 22: 175-81.
- Fraser F, Zasiadczyk L, Strzezek J (2010) Interactions of egg yolk lipoprotein fraction with boar spermatozoa assessed with a fluorescent membrane probe. Folia Histochemica et Cytobiologica 48: 292-298.
- Pillet E, Duchamp G, Batellier F, Beaumal V, Anton M, et al (2011) Egg yolk plasma can replace egg yolk in stallion freezing extenders. Theriogenology 75: 105-114. PMID: 20833417.
- Rodriguez H, Ohanian C, Bustos-Obregon E (1985) Nuclear chromatin decondensation of spermatozoa in vitro: a method for evaluating the fertilizing ability of ovine semen. Int J Androl 8:147-158.
- Simonsson A (1994) Näringsrekommendationer och fodermedelstabeller till svin [Nutrient and metabolizable energy recommendations for swine, in Swedish]. Swedish University of Agricultural Sciences, SLU Info. Rapporter, Husdjur.
- Tejerina F, Buranaamnuay K, Saravia F, Wallgren M, Rodriguez-Martinez H (2008) Assessment of motility of ejaculated, liquid-stored boar spermatozoa using computerized instruments. Theriogenology69: 1129-1138.
- Rodriguez-Martinez H, Wallgren M (2011) Advances in Boar Semen Cryopreservation. Vet Med Int pii: 396181.
- Peña F, Johannisson A, Wallgren M, Rodriguez-Martinez H (2004) Antioxidant supplementation of boar spermatozoa from different fractions of the ejaculate improves cryopreservation: changes in sperm membrane lipid architecture. Zygote12:117-124.
- Waterhouse K, Hofmo P, Tverdal A, Miller RJr (2006) Within and between breed differences in freezing tolerance and plasma membrane fatty acid composition of boar sperm. Reproduction131: 887-894.
- Aitken RJ (1995) Free radicals, lipid peroxidation and sperm function. Reprod Fertil Dev7: 659-668.
- Zhao YY, Xiong Y, Curtis JM (2011) Measurement of phospholipids by hydrophilic interaction liquid chromatography coupled to tandem mass spectrometry: The determination of choline containing compounds in foods. J Chromatogr A 1218: 5470-5479.
- Scholfield CR (1981) Composition of Soybean Lecithin. Journal of the American Oil Chemists' Society 58: 889-892.
- Jiang ZL, Li QW, Li WY, Hu JH, Zhao HW, et al. (2007) Effect of low density lipoprotein on DNA integrity of freezing-thawing boar sperm by neutral comet assay. Anim Reprod Sci 99: 401-407.
- Hu H, Li QW, Li G, Chen XY, Yang H, et al. (2006) The cryoprotective effect on frozen–thawed boar semen of egg yolk low density lipoproteins. Asian-Aust J Anim Sci 19: 486-490.
- Long JA, Conn TL (2012) Use of phosphatidylcholine to improve the function of turkey semen stored at 4°C for 24 hours. Poult Sci 91: 1990-1996.
- Najafi A, Daghigh-Kia H, Dodaran HV, MehdipourM, Alvarez-Rodriguez M (2017) Ethylene glycol, but not DMSO, could replace glycerol inclusion in soybean lecithin-based extenders in ram sperm cryopreservation. Anim Reprod Sci 177: 35-41.
- Flores E, Fernández-Novell JM, Peña A, Rigau T, Rodríguez-Gil JE (2010) Cryopreservation-induced alterations in boar spermatozoa mitochondrial function are related to changes in the expression and location of midpiece mitofusin-2 and actin network. Theriogenology 74: 354-363.
- King RJ, MacBeth MC (1981) Interaction of the lipid and protein components of pulmonary sulfactant role of phosphatidylglycerol and calcium. Biochimica et Biophysica Acta 647: 159-168.
- Fraser L, Strzezek J (2007) Effect of different procedures of ejaculate collection, extenders and packages on DNA integrity of boar spermatozoa following freezing-thawing. Anim Reprod Sci 99: 317-329.
- Brouwers JF, Silva PF, Gadella BM (2005) New assays for detection and localization of endogenous lipid peroxidation products in living boar sperm after BTS dilution or after freeze-thawing. Theriogenology 63: 458-469.
- Burh MM, Curtis EF, Kakuda NS (1994) Composition and behavior of head membrane lipids of fresh and cryopreserved boar sperm. Cryobiology 31: 224-238.
- Yeste M, Estrada E, Rivera Del A´lamo MM, Bonet, S Rigau T, et al (2014) The increase in phosphorylation levels of serine residues of protein HSP70 during holding time at 17 °C is concomitant with a higher cryotolerance of boar spermatozoa. PLoS One 9: e90887.
- Okazaki T, Abe S, Yoshida S, Shimada M (2009) Seminal plasma damages sperm during cryopreservation, but its presence during thawing improves semen quality and conception rates in boars with poor post-thaw semen quality. Theriogenology 71: 491-498.
- Fernández-Gago R, Álvarez-Rodríguez M, Alonso ME, González JR, Alegre B, et al. (2016) Thawing boar semen in the presence of seminal plasma improves motility, modifies subpopulation patterns and reduces chromatin alterations. Reprod. Fertil. Dev. doi: 10.1071/RD15530.
- Saravia F, Wallgren M, Johannisson A, Calvete JJ, Sanz L, et al (2009) Exposure to the seminal plasma of different portions of the boar ejaculate modulates the survival of spermatozoa cryopreserved in MiniFlatPacks, Theriogenology 71: 662-675.
- Rath D, Niemann H (1997) In vitro fertilization of porcine oocytes with fresh and frozen-thawed ejaculated or frozen-thawed epididymal semen obtained from identical boars. Theriogenology 47: 785-937.
- Rodriguez-Martinez H, Larsson B, Pertoft H (1997) Evaluation of sperm damage and techniques for sperm clean-up. Reprod Fertil Dev 9: 297-308.