Keywords
Diabetes mellitus; Pathogenesis; Suprachiasmatic nucleus; Cryptochrome
What is the biological clock?
In mammals, the components of the biological clock function peripherally in a complex network and modulate the transcription of specific target genes and their products, expressed rhythmically in a 24 h rhythm. The central biological clock includes the suprachiasmatic nucleus, which rhythmically 'tunes' peripheral clocks (Figure 1) [1-8].
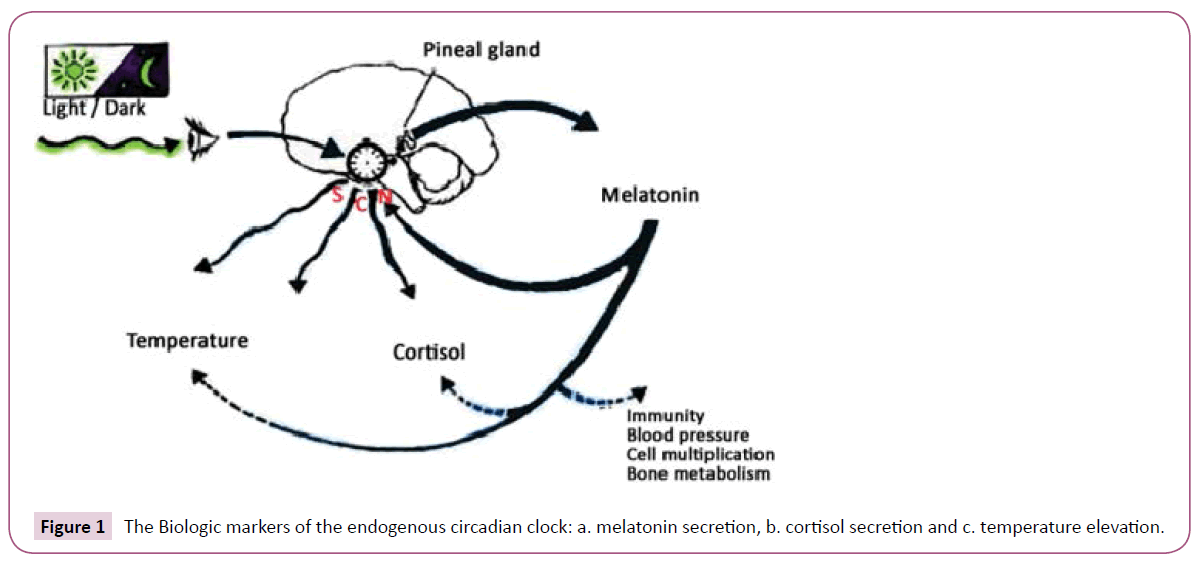
Figure 1: The Biologic markers of the endogenous circadian clock: a. melatonin secretion, b. cortisol secretion and c. temperature elevation.
Having the molecular basis of SCN being clarified the so-called core clock genes such as per1-2, cry1-2, bmal1 [9,10] and the demonstration that these genes are rhythmically expressed in peripheral tissues, including the liver, adrenal and pituitary glands [11-13], this central clock model is reshaped [14-16].
Clock/Bmal, formed by the product of the genes circadian locomotor output cycles kaput [Clock] and brain and muscle aryl hydrocarbon receptor nuclear translocator like-Arntl (Bmal) represents the transcription initiator of the feedback loops [17].
Clock/Bmal binds to E-box in the promoter regions of target genes Period homolog 1, 2 and 3 genes (Per1, Per2, Per3), Cryptochrome genes (Cry1, Cry2), retinoic acid-related orphan receptor (Rora, Rorb, Rorc) and Rev-Erb nuclear orphan receptor(Rev-Erbα, Rev- Erbβ) to activate their expression [18,19].
Transcription of Pers and Crys is initiated during the day, while degradation during the night. PER and CRY proteins enter the nucleus, probably as a multimeric complex (PER/CRY), and inhibit Clock/Bmal-mediated transcription [20,21].
As to the role of the PER/CRY complex, Cryptochrome (CRY) is a blue-light sensor, which regulates neuronal firing rate, that is circadian entrainment in Drosophila melanogaster [22]. Light activates CRY. When CRY is activated TIM [timeless] degrades, tuning the clock daily. In the absence of cry, the clock can still be activated by light, though the mechanisms are unclear; possibly, there are two types of clock neurons having differential sensitivities to light and temperature [23].
With regard to the remainder feedback loop, the ROR/Bmal/Rev- Erb (RBR) frames the original shape [20]. ROR acts as an activator of Bmal and Rev-Erb as an inhibitor which results in tuning of Bmal transcription [24,25]. Suppression of Rev-Erbα expression resulted in elevated Clock mRNA expression consistent with Rev- Erbα's role as a transcriptional repressor, adding a regulation role of the activity of the Bmal1/Clock heterodimer by regulation of the expression of both the Bmal1 and Clock genes directly (Figure 2) [26]. Other nuclear receptor family members, such as peroxisome proliferator activated receptors-α/γ, estrogen receptor-α, and retinoic acid receptor through their ligands link the clock mechanism with peripheral stimuli output [27].
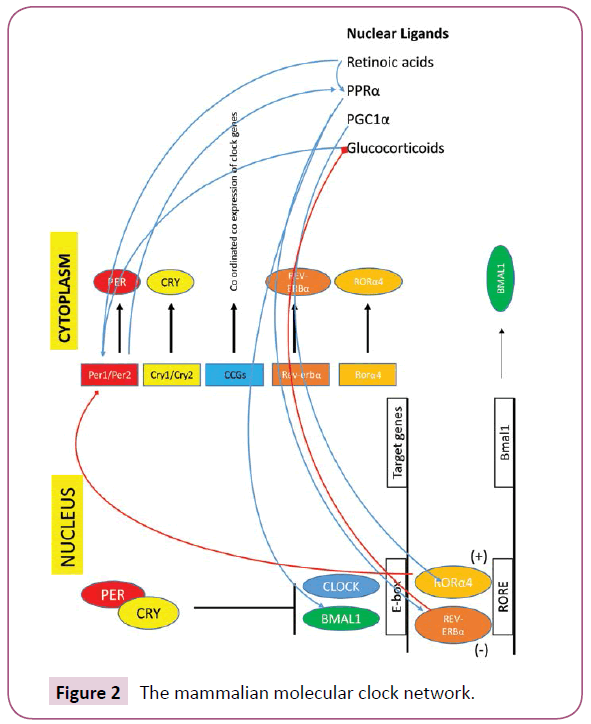
Figure 2: The mammalian molecular clock network.
Circadian entrainment and metabolism
Secondary peripheral clocks are found in various tissues, such as the liver, pancreas, and adipose tissue. These clocks control the rhythmic patterns of metabolic processes. If the function of the central or peripheral clocks is disturbed diabetes mellitus or obesity emerges [28]. For instance, Lamia et al. [29] have described the metabolic phenotype of total vs. liver-specific Bmal1-deficient mice. The latter present with hypoglycemia, in the presence of an intact central biological clock mechanism, emphasizing the importance of the function of peripheral clocks for the homeostasis of glucose.
Circadian Rhythms and Cardiovascular Disease
Circadian expression of clock genes was reported in cardiovascular tissues by several investigators. The measurement of core clock gene expression revealed rhythmicity in the heart, aorta, and kidney [30,31]. Almost all of the known core clock genes, such as clock, bmal1, cry1/2, and per1/2/3 genes, are expressed in the heart [32]. Expression of some genes in the heart oscillates with circadian rhythmicity. Regulation of heart's systolic function, oxygen metabolism and resistance to ischaemic disorders is done by the clock seated in the cardiomyocyte. These same biological processes are also directly influenced i.e., by modification of proteins by monosaccharides of O−linked β− N−acetylglucosamine (O−GlcNAc) [33]. Despite of the integral relation between clocks and metabolism, little is known regarding the direct influence of a peripheral clock on cellular responses to fatty acids. Tsai et al. [34] to address this issue utilized a genetic model of disrupted clock function specifically in cardiomyocytes in vivo (termed cardiomyocyte clock mutant). There are data that the cardiomyocyte clock modulates triglyceride metabolism in the myocardiocyte [34].
Further, Rev-Erbα is present in vascular wall cells including macrophages [35-37]. Rev-Erbα downregulates toll like receptor, thus inhibiting the inflammatory cascade. These data suggest a positive impact of Rev-Erbα on atherosclerotic lesions. Likewise, Rev-Erbα has been identified as a potent negative regulator of plasminogen activator inhibitor (PAI)-1, an important inhibitor of the fibrinolysis cascade that may promote the development of atherothrombosis [38]. In vitro results suggest a regulatory link between Rev-Erbα and the NF−kappaB pathway in vascular wall cells, too [37]. Rev-Erbα, besides exerting a regulatory role on lipid metabolism, adipogenesis and vascular inflammation, also cross talks with several other nuclear receptors involved in energy homeostasis. Therefore this nuclear orphan receptor by coupling metabolic and circadian signals [38,39], gives a time cue to metabolism−becoming so a potential modulator of the morning susceptibility to myocardial infarction [40].
The transcript levels of potassium channels Kv1.5 and Kv4.2 also indicate apparent circadian fluctuation [41]. The circadian variation of the onset of cardiac arrhythmia might be attributed to intrinsic circadian rhythms in potassium channel expression. Although the exact mechanisms regulating the circadian expression of these genes are unknown, some of these genes may be clock output genes in the heart. In the heart with hypertrophy in rat’s experiments [42], the tuned transcription of anp is inhibited along with hlf and dbp. This reduction of circadian expression of anp may perturb the adaptive response of cardiomyocytes to external stimuli and accelerate the hypertrophic processes. The circadian rhythm of mean blood pressure [MBP] was selectively disrupted in sinoaortic denervated rats under 12 h light−dark cycles as a result of an increase in MBP during the light period and disappeared under constant darkness [43].
Regarding the clinical setting, Manios et al. [44] recently evaluated the association between the rate of blood pressure variation, derived from ambulatory blood pressure monitoring data analysis, and the severity and topography of coronary artery lesion in a cohort of normotensive patients with suspected coronary artery disease. Abrupt variations in blood pressure are related to atherosclerosis and enhance cardiac hypertrophy [45].
A group from Amsterdam had already determined the continuous 24 h profile of mean arterial pressure, heart rate, stroke volume, cardiac output, and total peripheral resistance in eight healthy ambulatory volunteers. During the night time, compared with the daytime average, there was a decrease in blood pressure (9 mm Hg), heart rate (18 beats per minute), and cardiac output (29%); stroke volume showed a small decrease [7%], and total peripheral resistance increased unexpectedly by 22% [46].
Kollias et al. [47] earlier demonstrated a circadian variation of endothelial function and arterial stiffness in hypertension, that is endothelial function is decreased in the early morning in hypertensive patients, whereas arterial stiffness is increased in the evening. No ulterior, Zakopoulos et al. [48] presented that diurnal variations of continuously monitored interstitial glucose concentrations significantly associate with blood pressure levels in both normotensive and hypertensive humans, indicating a common pathway of circadian autoregulation, probably stemming from both central mechanisms and peripheral inputs which in turn exert a feedback loop.
Actually, the amplitude of this matter disaggregated by Guo et al., in American Heart Journal in 2003. Results of these studies confirm that most cardiovascular physiological parameters (such as heart rate, blood pressure, electrocardiogram indices) and pathophysiological events [myocardial ischemia/infarction, sudden cardiac death) show circadian rhythms [49]. Interestingly, Mann et al. had prior showed that patients with primary autonomic failure and very low sympathetic and parasympathetic activities also have a high incidence of nondipping [50], suggesting that it is the inability to modulate autonomic tone that is responsible for the nondipping phenomenon [51]. Other factors be taken into account, a quantitative analysis of the effects of activity and time of day on the diurnal variations of blood pressure clearly indicated there was no significant diurnal variation of BP which is independent of activity [52]. To a deeper approach, in normoalbuminuric normotensive type 1 diabetic patients without any degree of autonomic dysfunction, according to traditional cardiovascular tests, diastolic BP night/day ratio is associated with an increased glomerular filtration rate and an increased extracellular volume. The disruption of the circadian rhythm of sympathovagal activity in non−dipper patients was associated with higher levels in systolic and diastolic blood pressure and with a reduced decline in their levels during the night [53]. Dippers suffering from hypertension independently present with cardiovascular events without having a positive history [54].
Is there a clinical relevance of the biologic clock to pregnancy? The endogenous placental clock: Circadian variation of the HPA axis is dependent on clock gene rhythms in the hypothalamus, but it is not known whether pregnancy-induced changes in maternal glucocorticoid levels are mediated via this central clock [55]. Does a placenta function like a tuned clock? Recent recognition that clock genes are expressed in the placenta, together with observations linking circadian disruption with compromised placental function, suggests that circadian variation may be an important component of the normal placental phenotype [56]. Of note is that, for both pregnant women with complications in either the second or third trimester and healthy pregnant women, melatonin levels were the highest at night, decreased throughout the early morning, and were the lowest during the day, demonstrating significant daily fluctuations [57].
In fact, maternal melatonin enters the fetal circulation with ease providing photoperiodic information to the fetus. Melatonin protects placenta from maternal oxidative stress, modulates endocrine function and tunes the function of immunocompetent cells [58]. Melatonin reduces oxidative damage in the placenta and may improve hemodynamics and nutrient transfer at the placental-uterine interface [59].
Pérez et al. has shown the existence of a biological clock in this tissue, by studying the expression of circadian locomotors output cycles kaput [Clock], brain and muscle arnt-like (Bmal)1, period(Per)2, and cryptochrome (Cry)1 mRNAs at 00, 04, 08, 12, 16, and 20 h by qPCR. The clock genes are present in human placenta, reinforcing the hypothesis of the placental biological clock [60]. Moreover, alterations in the hypothalamic pituitary adrenal axis during pregnancy and the placental clock determine the length of parturition [61].
Is a Dysfunction of the Placental Clock Related to Preeclampsia?
Maternal oxidative stress predisposes to preeclampsia. Simultaneously, melatonin, as a seasonal factor, has been suggested to be involved in a successful pregnancy [62-64]. Το be brought to safe conclusions we should carefully consider the pathophysiologic process taking place in preeclampsia (Figure 3). Κarteris showed that placental CRH-R1 alpha mRNA levels (as shown by quantitative RT-PCR) and protein levels [shown by Western blotting analysis] were significantly reduced in all of the complicated pregnancies [65]. Previously, Karteris working team supported that there is a CRH placental clock that is active from the early stages of pregnancy and determines the length of gestation and the timing of parturition. CRH can influence human reproductive tissue function via specific CRH receptors. Two distinct CRH receptors have been cloned (R1 and R2) that exist as two alternatively spliced forms (alpha and beta) [66]. Actually, CRH participates in the pathophysiology of pre-eclampsia through the induction of macrophage-mediated apoptosis of extravillous trophoblasts (Figure 4).
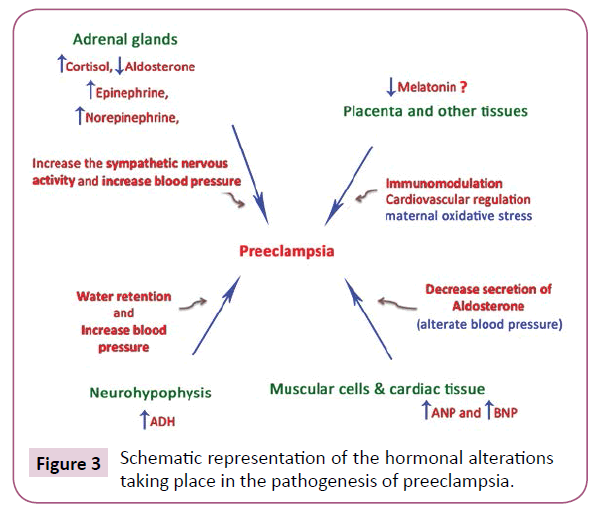
Figure 3: Schematic representation of the hormonal alterations taking place in the pathogenesis of preeclampsia.
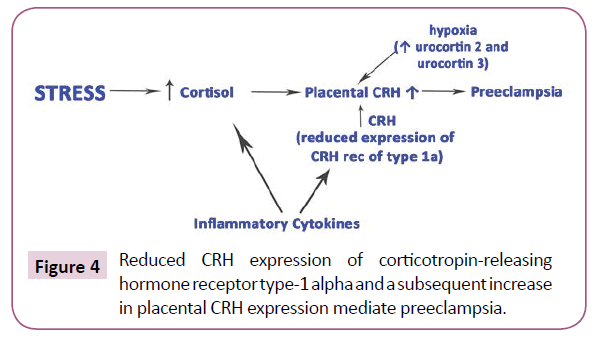
Figure 4: Reduced CRH expression of corticotropin-releasing hormone receptor type-1 alpha and a subsequent increase in placental CRH expression mediate preeclampsia.
It was found that the expression of CRH was increased in trophoblasts of the placental bed biopsy specimens from preeclamptic pregnancies [67]. Nevertheless, a question mark remains as to the role of melatonin in the pathophysiology of preeclampsia and the associated input driven by the biological clock.
Shimada showed that pregnant women with hypertension or diabetes had lower levels of salivary melatonin during the day than healthy pregnant women [57]. Moreover, Reiter suggested that due to the potent antioxidant actions of melatonin, coupled with its virtual absence of toxicity, this indoleamine may have utility in the treatment of pre-eclampsia, intrauterine growth restriction, placental and fetal ischemia/reperfusion [68].
Clinical approach of vascular disease including the time of pregnancy
Physicians, care managers, and patients agreed regarding the positive impact on patient health and self-management, and attributed the outcomes to the strong “partnership” between the care manager and the patient and the collaboration between the physician and the care manager [69]. Previously, it was confirmed by Ciccone et al. that carotid atherosclerosis is associated with coronary atherosclerosis, highlighting the importance of screening for ischemic heart disease in patients with asymptomatic carotid plaques, considering eventually plaque morphology[symmetry, composition, eccentricity or concentricity of the plaque] for patient stratification [70]. Moreover, considering the disturbed function of the placental clock and preterm delivery, different functional cardiac characteristics have been observed in term and preterm neonates by echocardiography and tissue doppler imaging, while increased size of the left ventricle and altered left ventricular diastolic function was found in children born very low birth weight preterm [71,72]. What do we observe? Certainly, atherosclerosis and ischemia are common manifestations of a disorder, which at least initially involves unwound clock function. Further, cardiovascular disease of the mother or reasons such as lower placental perfusion, increased blood pressure in the mother and preeclampsia, often resulting in intrauterine growth restriction (IUGR), perpetuate vascular disease. That is because developmental cardiovascular alterations and potential mechanisms link preterm birth to the risk of hypertension and cardiovascular diseases into adulthood [73]. Of note is that, impairments to vascular system development may initially occur before preterm birth via IUGR and exposure to an inflammatory and antiangiogenic environment [74].
Conclusion
Given the broaden role of peripheral circadian loops in lipid metabolism affecting the development of atherosclerosis and vascular wall pathophysiology or autonomic tone and blood pressure per se, regulation of the expression of both the Bmal1 and Clock genes represents a promising target for the treatment of cardiovascular abnormalities resulting from chronic aberrant circadian rhythms. Perhaps, treatment with melatonin is the appropriate therapy for preeclampsia.
References
- Panda S, Hogenesch JB, Kay SA (2002) Circadian rhythms from flies to human. Nature 417: 329-335.
- Mohawk JA, Green CB, Takahashi JS (2012) Central and peripheral circadian clocks in mammals. Ann Rev Neurosci 35: 445-462.
- Gale JE, Cox HI, Qian J, Block GD, Colwell CS, et al. (2011) Disruption of circadian rhythms accelerates development of diabetes through pancreatic beta-cell loss and dysfunction. J Biol Rhythms 26: 423-433.
- Krystal AD (2012) How the circadian rhythm affects sleep, wakefulness, and overall health: background for understanding shift work disorder. J Clin Psychiatry 73: e05.
- Yang Z, Zhang W, Wang M, Ruan D, Chen J (2012) Effects of daytime, night and sleep pressure on long-term depression in the hippocampus in vivo. Neurosci Lett 511: 106-109.
- Jeyaraj D, Haldar SM, Wan X, McCauley MD, Ripperger JA, et al. (2012) Circadian rhythms govern cardiac repolarization and arrhythmogenesis. Nature 483: 96-99.
- Greene MW (2012) Circadian rhythms and tumor growth. Cancer Lett 318: 115-123.
- Thoennissen NH, Thoennissen GB, Abbassi S, Nabavi-Nouis S, Sauer T, et al. (2012) Transcription factor CCAAT/enhancer-binding protein alpha and critical circadian clock downstream target gene PER2 are highly deregulated in diffuse large B-cell lymphoma. Leuk Lymphoma 53: 1577-1585.
- Doherty CJ, Kay SA (2010) Circadian control of global gene expression patterns. Ann Rev Genet 44: 419-444.
- Ukai H, Ueda HR (2010) Systems biology of mammalian circadian clocks. Ann Rev Physiol 72: 579-603.
- Cailotto C, Lei J, van der Vliet J, van Heijningen C, van Eden CG, et al. (2009) Effects of nocturnal light on (clock) gene expression in peripheral organs: a role for the autonomic innervation of the liver. PLoS One 4: e5650.
- Fahrenkrug J, Hannibal J, Georg B (2008) Diurnal rhythmicity of the canonical clock genes Per1, Per2 and Bmal1 in the rat adrenal gland is unaltered after hypophysectomy. J Neuroendocrinol 20: 323-329.
- Kalsbeek A, van der Spek R, Lei J, Endert E, Buijs RM, et al. (2012) Circadian rhythms in the hypothalamo-pituitary-adrenal (HPA) axis. Mol Cell Endocrinol 349: 20-29.
- Kaeffer B, Pardini L (2005) Clock genes of Mammalian cells: practical implications in tissue culture. In vitro Cell Dev Biol Anim 41: 311-320.
- Saini C, Suter DM, Liani A, Gos P, Schibler U (2011) The mammalian circadian timing system: synchronization of peripheral clocks. Cold Spring Harb Symp Quant Biol 76: 39-47.
- Chen Z, Yoo SH, Park YS, Kim KH, Wei S, et al. (2012) Identification of diverse modulators of central and peripheral circadian clocks by high-throughput chemical screening. Proc Natl Acad Sci U S A 109: 101-106.
- Ko CH, Takahashi JS (2006) Molecular components of the mammalian circadian clock. Hum Mol Genet 2: 271-277.
- Reppert SM, Weaver DR (2002) Coordination of circadian timing in mammals. Nature 418: 935-941.
- Ueda HR, Chen W, Adachi A, Wakamatsu H, Hayashi S, et al. (2002) A transcription factor response element for gene expression during circadian night. Nature 418: 534-539.
- Zhang EE, Kay SA (2010) Clocks not winding down: unravelling circadian networks. Nat Rev Mol Cell Biol 11: 764-776.
- Lee C, Etchegaray JP, Cagampang FR, Loudon AS, Reppert SM (2001) Posttranslational mechanisms regulate the mammalian circadian clock. Cell 107: 855-867.
- Fogle KJ, Parson KG, Dahm NA, Holmes TC (2011) Cryptochrome is a blue-light sensor that regulates neuronal firing rate. Science 331: 1409-1413.
- Yoshii T, Hermann C, Helfrich-Förster C (2010) Cryptochrome-positive and -negative clock neurons in Drosophila entrain differentially to light and temperature. J Biol Rhythms 25: 387-398.
- Rush BL, Murad A, Emery P, Giebultowicz JM (2006) Ectopic cryptochrome renders TIM light sensitive in the Drosophila ovary. J Biol Rhythms 21: 272-278.
- Guillaumond F, Dardente H, Giguère V, Cermakian N (2005) Differential control of Bmal1 circadian transcription by REV-ERB and ROR nuclear receptors. J Biol Rhythms 20: 391-403.
- Crumbley C, Burris TP (2011) Direct regulation of CLOCK expression by REV-ERB. PLoS One 6: e17290.
- Teboul M, Gréchez-Cassiau A, Guillaumond F, Delaunay F (2009) How nuclear receptors tell time. J Appl Physiol 107: 1965-1971.
- Mazzoccoli G, Pazienza V, Vinciguerra M (2012) Clock genes and clock-controlled genes in the regulation of metabolic rhythms. Chronobiol Int 29: 227-251.
- Lamia KA, Storch KF, Weitz CJ (2008) Physiological significance of a peripheral tissue circadian clock. Proc Natl Acad Sci USA 105: 15172-15177.
- Nonaka H, Emoto N, Ikeda K, Fukuya H, Rohman MS, et al. (2001)Angiotensin II induces circadian gene expression of clock genes in cultured vascular smooth muscle cells. Circulation 104: 1746-1748.
- Maemura K, de la Monte SM, Chin MT, Layne MD, Hsieh CM, et al. (2000) CLIF, a novel cycle-like factor, regulates the circadian oscillation of plasminogen activator inhibitor-1 gene expression. J Biol Chem 275: 36847-36851.
- Young ME (1985) Anticipating anticipation: pursuing identification of cardiomyocyte circadian clock function. J Appl Physiol 107: 1339-1347.
- Durgan DJ, Pat BM, Laczy B, Bradley JA, Tsai JY, et al. (2011) O-GlcNAcylation, novel post-translational modification linking myocardial metabolism and cardiomyocyte circadian clock. J Biol Chem 286: 44606-44619.
- Tsai JY, Kienesberger PC, Pulinilkunnil T, Sailors MH, Durgan DJ, et al. (2010) Direct regulation of myocardial triglyceride metabolism by the cardiomyocyte circadian clock. J Biol Chem 285: 2918-2929.
- Raspé E, Duez H, Mansén A, Fontaine C, Fiévet C, et al. (2002) Identification of Rev-erbalpha as a physiological repressor of apoC-III gene transcription. J Lipid Res 43: 2172-2179.
- Fontaine C, Rigamonti E, Pourcet B, Duez H, Duhem C, et al. (2008)The nuclear receptor Rev-erbalpha is a liver X receptor (LXR) target gene driving a negative feedback loop on select LXR induced pathways in human macrophages. Mol Endocrinol 22: 1797-1811.
- Migita H, Morser J, Kawai K (2004) Rev-erbalpha upregulates NF-kappaB-responsive genes in vascular smooth muscle cells. FEBS Lett 561: 69-74.
- Wang J, Yin L, Lazar MA (2006) The orphan nuclear receptor Rev-erb alpha regulates circadian expression of plasminogen activator inhibitor type 1. J Biol Chem 281: 33842-33848.
- Duez H, Staels B (2008) Rev-erb alpha gives a time cue to metabolism. FEBS Lett 582: 19-25.
- Young ME, Wilson CR, Razeghi P, Guthrie PH, Taegtmeyer H (2002) Alterations of the circadian clock in the heart by streptozotocin-induced diabetes. J Mol Cell Cardiol 34: 223-231.
- Yamashita T, Sekiguchi A, Iwasaki YK, Sagara K, Hatano S, et al. (2005) Cibenzoline attenuates upregulation of Kv1.5 channel gene expression by experimental paroxysmal atrial fibrillation. Int Heart J 46: 279-288.
- Young ME, Razeghi P, Taegtmeyer H (2001) Clock genes in the heart: characterization and attenuation with hypertrophy. Circ Res 88: 1142-1150.
- Makino M, Hayashi H, Takezawa H, Hirai M, Saito H, et al. (1997) Circadian rhythms of cardiovascular functions are modulated by the baroreflex and the autonomic nervous system in the rat. Circulation 96: 1667-1674.
- Manios E, Stamatelopoulos K, Tsivgoulis G, Barlas G, Koroboki E, et al. (2011) Time rate of blood pressure variation: a new factor associated with coronary atherosclerosis. J Hypertens 29: 1109-1114.
- Zakopoulos NA, Tsivgoulis G, Barlas G, Spengos K, Manios E, et al. (2006) Impact of the time rate of blood pressure variation on left ventricular mass. J Hypertens 24: 2071-2077.
- Veerman DP, Imholz BP, Wieling W, Wesseling KH, van Montfrans GA (1995) Circadian profile of systemic hemodynamics. Hypertension 26: 55-59.
- Kollias GE, Stamatelopoulos KS, Papaioannou TG, Zakopoulos NA, Alevizaki M, et al. (2009) Diurnal variation of endothelial function and arterial stiffness in hypertension. J Hum Hypertens 23: 597-604.
- Zakopoulos NA, Dolianitis KJ, Theodorakis MJ, Manios ED, Alevizaki M, et al. (2008) Diurnal correlation of ambulatory blood pressure and interstitial glucose in patients with normal glucose tolerance. Blood Press Monit 13: 309-317.
- Guo YF, Stein PK (2003) Circadian rhythm in the cardiovascular system: chronocardiology. Am Heart J 145: 779-786.
- Mann S, Altman DG, Raftery EB, Bannister R (1983) Circadian variation of blood pressure in autonomic failure. Circulation 68: 477-483.
- Biaggioni I (2008) Circadian clocks, autonomic rhythms, and blood pressure dipping. Hypertension 52: 797-798.
- Clark LA, Denby L, Pregibon D, Harshfield GA, Pickering TG, et al. (1987) A quantitative analysis of the effects of activity and time of day on the diurnal variations of blood pressure. J Chronic Dis 40: 671-681.
- Izzedine H, Launay-Vacher V, Deray G (2006) Abnormal blood pressure circadian rhythm: a target organ damage? Int J Cardiol 107: 343-349.
- Fagard RH, Thijs L, Staessen JA, Clement DL, et al. (2009) Night-day blood pressure ratio and dipping pattern as predictors of death and cardiovascular events in hypertension. J Hum Hypertens 23: 645-653.
- Wharfe MD, Mark PJ, Wyrwoll CS, Smith JT, Yap C, et al. (2016) Pregnancy-induced adaptations of the central circadian clock and maternal glucocorticoids. J Endocrinol 228: 135-147.
- Waddell BJ, Wharfe MD, Crew RC, Mark PJ (2012) A rhythmic placenta? Circadian variation, clock genes and placental function. Placenta 33: 533-539.
- Shimada M, Seki H, Samejima M, Hayase M, Shirai F (2016) Salivary melatonin levels and sleep-wake rhythms in pregnant women with hypertensive and glucose metabolic disorders: A prospective analysis. Biosci Trends 10: 34-41.
- Tamura H, Nakamura Y, Terron MP, Flores LJ, Manchester LC, et al. (2008) Melatonin and pregnancy in the human. Reprod Toxicol 25: 291-303.
- Reiter RJ, Tan DX, Tamura H, Cruz MH, Fuentes-Broto L (2014) Clinical relevance of melatonin in ovarian and placental physiology: a review. Gynecol Endocrinol 30: 83-89.
- Pérez S, Murias L, Fernández-Plaza C, Díaz I, González C, et al. (2015) Evidence for clock genes circadian rhythms in human full-term placenta. Syst Biol Reprod Med 61: 360-366.
- Smith R (1998) Alterations in the hypothalamic pituitary adrenal axis during pregnancy and the placental clock that determines the length of parturition. J Reprod Immunol 39: 215-220.
- Lanoix D, Beghdadi H, Lafond J, Vaillancourt C (2008) Human placental trophoblasts synthesize melatonin and express its receptors. J Pineal Res 45: 50-60.
- Soliman A, Lacasse AA, Lanoix D, Sagrillo-Fagundes L, Boulard V, et al. (2015) Placental melatonin system is present throughout pregnancy and regulates villous trophoblast differentiation. J Pineal Res 59: 38-46.
- Zeng K, Gao Y, Wan J, Tong M, Lee AC, et al. (2016) The reduction in circulating levels of melatonin may be associated with the development of preeclampsia. J Hum Hypertens 30: 666-671.
- Karteris E, Goumenou A, Koumantakis E, Hillhouse EW, Grammatopoulos DK (2003) Reduced expression of corticotropin-releasing hormone receptor type-1 alpha in human preeclamptic and growth-restricted placentas. J Clin Endocrinol Metab 88: 363-370.
- Karteris E, Grammatopoulos D, Dai Y, Olah KB, Ghobara TB, et al. (1998) The human placenta and fetal membranes express the corticotropin-releasing hormone receptor 1alpha (CRH-1alpha) and the CRH-C variant receptor. J Clin Endocrinol Metab 83: 1376-1379.
- Petsas G, Jeschke U, Richter DU, Minas V, Hammer A, et al. (2012) Aberrant expression of corticotropin-releasing hormone in pre-eclampsia induces expression of FasL in maternal macrophages and extravillous trophoblast apoptosis. Mol Hum Reprod 18: 535-45.
- Reiter RJ, Tan DX, Korkmaz A, Rosales-Corral SA (2014) Melatonin and stable circadian rhythms optimize maternal, placental and fetal physiology. Hum Reprod Update 20: 293-307.
- Ciccone MM, Aquilino A, Cortese F, Scicchitano P, Sassara M, et al. (2010) Feasibility and effectiveness of a disease and care management model in the primary health care system for patients with heart failure and diabetes (Project Leonardo). Vasc Health Risk Manag 6: 297-305.
- Ciccone MM, Niccoli-Asabella A, Scicchitano P, Gesualdo M, Notaristefano A et al. (2011) Cardiovascular risk evaluation and prevalence of silent myocardial ischemia in subjects with asymptomatic carotid artery disease. Vasc Health Risk Manag 7: 129-134.
- Ciccone MM, Scicchitano P, Zito A, Gesualdo M, Sassara M, et al. (2011) Different functional cardiac characteristics observed in term/preterm neonates by echocardiography and tissue doppler imaging. Early Hum Dev 87: 555-558.
- Ciccone MM, Cortese F, Gesualdo M, DI Mauro A, Tafuri S, et al. (2016) The role of very low birth weight and prematurity on cardiovascular disease risk and on kidney development in children: a pilot study. Minerva Pediatr.
- Bertagnolli M, Luu TM, Lewandowski AJ, Leeson P, Nuyt AM, et al. (2016) Preterm Birth and Hypertension: Is There a Link? Curr Hypertens Rep 18: 28.
- Tsao PN, Wei SC, Su YN, Chou HC, Chen CY, et al. (2005) Excess soluble fms-like tyrosine kinase 1 and low platelet counts in premature neonates of preeclamptic mothers. Pediatrics 116: 468-472.

