Samad Javadi1,2, HojjatAllah Alaei3*, Seyid Ebrahim Hosseini2* and Mohammad Amin Edalatmanesh2
1Department of Biology, College of Science, Fars Science and Research Branch, Islamic Azad University, Fars, Iran
2Department of Biology, College of Science, Shiraz Branch, Islamic Azad University, Shiraz, Iran
3Department of Physiology, School of Medicine, Isfahan University of Medical Sciences, Isfahan, Iran
*Corresponding Authors:
HojjatAllah Alaei
Department of Physiology, School of Medicine
Isfahan University of Medical Sciences, Isfahan, Iran
Tel: +98 3137922407
Fax: +98 3136687898
E-mail: alaei@med.mui.ac.ir
Seyeid Ebrahim Hosseini
Department of Biology, College of Science
Shiraz Branch, Islamic Azad University, Shiraz, Iran
E-mail: ebrahim.hossini@yahoo.com
Received date: March 01, 2017; Accepted date: March 20, 2017; Published date: March 26, 2017
Citation: Javadi S, Alaei HA, Hosseini SE, Edalatmanesh MA (2017) The Effect of Glutamate AMPA Receptors Blockade into the Prelimbic on Morphine-induced CPP in Rat. J Addict Behav Ther 1:4.
Copyright: © 2017 Javadi S, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Keywords
PL; CPP; Addiction; CNQX; Morphine; AMPA
Introduction
Addiction is a growing problem in public health. Evidences have demonstrated that learning and memory play an important role in the development of opiate tolerance and dependence [1]. Many studies suggested that using morphine to relieve the pain, causes physical and psychological dependence that is recognized by the expression of withdrawal symptoms even after cessation of drug. [2]. Many parts of the brain involved in addiction disorder. The prelimbic area (PL) is a subregion of the prefrontal cortex and a critical component in the circuitry for drug-seeking behavior and addiction process [3-5].
Recent studies have revealed that the PL plays a crucial role in social cognition and emotional expression including response to rewards and punishment [6]. Pharmacological and anatomical findings suggested that glutamate is the excitatory neurotransmitter in the central nervous system and any abnormality in glutamate transmission may be associated with the pathophysiology of some disorders [7]. There is now compelling evidence that the glutamate plays a pivotal role in drug addiction [8]. It has been demonstrated that the glutamate system also controls opiate addiction via the α-amino-3- hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) receptors. The AMPA receptors have key role in drug-seeking [9] and addiction [10].
Abuse drugs alter expression of AMPA receptors in the nucleus accumbens. NAc is a key component of brain reward systems [11]. Previous findings suggested a critical role for intra- PL AMPA receptor transmission in the processing of opiate reward signaling [12] as, morphine stimulates glutamate release in the ventral tegmental area (VTA) via AMPA receptors [10]. Also behavioral studies have corroborated the importance of glutamatergic transmission through the AMPA receptors in the processing and recalling of drug-related associative cues [13] but, the role of the PL and its glutamatergic system in reward is not clear for all drugs especially morphine.
Therefore, we evaluated a series of behavioral studies to investigate the effect of intra-PL microinjection of the AMPA antagonist on the effective and the ineffective doses of morphine-induced CPP.
Methods
Animals
Our experiments were done on Male Wistar rats (weighing 250-300 g at the beginning of the experiment) provided by the Institute of Pasteur, Iran. They were housed 5/cage, with the room temperature maintained at 22 ± 2°C, under a relative humidity of 40%, and a 12:12 h light-dark cycle [14]. The experimental procedures were approved by the Committee on Animal Care and use of Isfahan University of Medical Sciences.
Drug
Morphine was prepared from Temad, Tehran (Iran) and the 6- cyano-7-nitroquinoxaline-2,3-Dione (CNQX) was bought from Sigma, (U.S.A.). Morphine dissolved in saline [14] and CNQX in saline 50%/DMSO 50% [15].
Surgical procedures
All rats were anesthetized with chloral hydrate, injected intraperitoneally (400 mg/kg). After shaving, their heads were located in a stereotaxic instrument; a cannula (22 G) was then implanted into the PL (AP=3.2 mm, L=0.7 mm, DV=3 mm), according to the point was defined by Paxinus and Watson [16]. Finally, the cannula was anchored in the skull by dental cement. In order to preempt infection, Gentamycin (40 mg/ml, i.p.) was administered immediately following the surgery.
Microinjection method
All animals were allowed to being recovered for 1 week before the beginning of the behavioral test [17-21] and they received the CNQX for the CPP experiment, before every morphine conditioning session. Animals were tested in a morphine-free state [19].
Initially, the rats were kept in hand and the injection needle related to the Hamilton syringe was placed in a short polyethylene tube (PE20) in the cannula. The microinjections were performed unilaterally, through lowering a stainless steel injector cannula with a length of 1 mm longer than the guide cannula into the PL. The injector cannula was connected to Hamilton syringe by polyethylene tube; next, different doses of the CNQX (0.5 μg/0.5 μl and 2.5 μg/0.5 μl) were injected with rate of 2 μl/min into the PL, 5 min before the acquisition/ expression phases of the experiment [20]. The injection cannula was left in the guide cannula for an additional 60 sec to facilitate diffusion of the drug and then removed [19-21]. Different doses of the CNQX were injected in the distinct animals and each animal only once was tested in the CPP procedure.
Apparatus
The CPP apparatus consists of three chambers (A, B, and C). Two chambers (A and B) are similar in size but different in color. The walls and floor of chamber A are black and white while the walls and floor of chamber B are all white. Chamber C is smaller and is connected to A and B chambers via a guillotine door [22].
Behavioral Testing-Place preference paradigm
The behavioral procedure of the CPP is done in five continuous days and has three distinct phases [18,22]. Among the different behavioral methods were used to measure drug reward in laboratory animals, conditioned place preference has been one of the most popular [23].
Preconditioning
In the pre-test (day 1) each rat was placed in chamber C while the middle door was opened and the rat was allowed free movement for 15 minutes in all chambers. The time spent in each chamber (A and B) was recorded by the apparatus.
Conditioning
This phase consists of 3 days (from day 2 to day 4) each lasting 45 minutes. Guillotine door was closed and daily injection was performed in two stages with a 6 h interval.
On the second day, after the subcutaneous injection of morphine, the rats were confined to one chamber of the apparatus for 45 minutes. Then with an interval of 6 h, following the injection of saline instead of morphine, the rats were confined to another side of the apparatus for 45 minutes.
On the third day, morphine and saline injections were contrary to day 2, while On day 4, morphine and saline injections were the same as day 2 [18,22].
Post conditioning
In this phase (day 5) the guillotine door was opened (as in the first day) and the rats could move freely in all chambers for 15 minutes; the time the morphine-injected rats spent in the chamber was recorded. The preferences were computed as the difference (in second) between the time spent in the morphine receiving chamber on the test day and the first day [18,22].
The effect of the CNQX on drug-naïve and morphine-conditioned animals
CPP difference score was calculated by subtracting the amount of time spent in the saline-paired compartment from the amount of time spent in the CNQX-paired compartment during the CPP test [13,24,25]. In another set of our experiments we tested, intra-PL microinjection different doses of CNQX versus intra-PL saline in drug-naïve animals [13].
Locomotor activity
In our experiment, the effect of intra-PL CNQX with administration of different doses of morphine on locomotor activity was measured. Locomotor activity index was calculated based on the numbers of compartment crossing in morphineinduced CPP [18].
Experimental Design
Morphine dose-response on the CPP paradigm
The effect of different doses of morphine (0.5, 1, 2.5, 5, 7.5 and 10 mg/kg) was tested on the CPP paradigm. Animals were divided into 6 groups, each including 7 rats; 5 groups received different doses of morphine while one group received saline. Morphine or saline was injected according to the CPP paradigm and then calculated the changes of preference.
Microinjection of the CNQX into the PL with different doses of morphine
According to the experiment design, the animals were divided into three surgical groups: for each dose of morphine in each phase of the CPP, therefore we used 12 groups (7 rats in each group). Sham group received intra-PL saline and morphine via intraperitoneal (i.p) without any microinjection of the CNQX while test groups received intra-PL low or high doses of CNQX before injection of morphine on days 2, 3 and 4 during the acquisition phase of the CPP paradigm and only received the CNQX in the post conditioning of the expression phase.
Histology
At the end of the our experiments, the rats were anesthetized and were perfused with 0.9% saline followed by 10% formalin; their brains were removed carefully and were placed in 10% formalin for 72 h before to being sliced. In order to evaluate the place of the antagonist injection and cannula, the PL sections were checked (Figure 1).
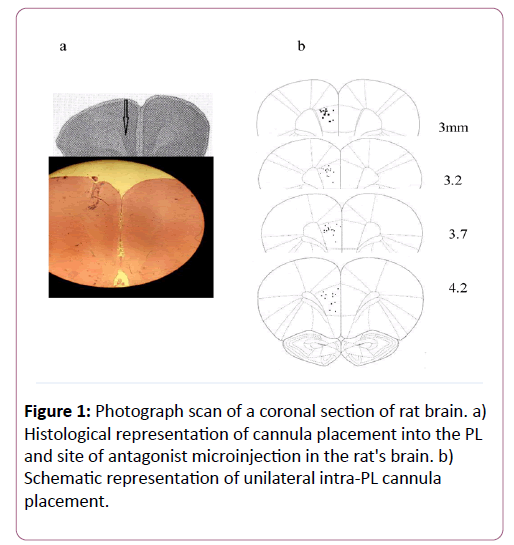
Figure 1: Photograph scan of a coronal section of rat brain. a) Histological representation of cannula placement into the PL and site of antagonist microinjection in the rat's brain. b) Schematic representation of unilateral intra-PL cannula placement.
Statistical analysis
Data were analyzed by the commercially available SPSS 22 software. The means of the groups were compared with one-way ANOVA followed by Tukey’s test. Results are presented as mean ± s.e.m and the difference (with p<0.05) between the experimental groups was considered as statically significant.
Results
Determining the effective dose of morphine-induced place preference
For determining the effective dose of morphine, saline and different doses of morphine were tested according to the CPP paradigm. Our findings showed that saline and low doses of morphine (0.5 and 1 mg/kg) did not induce place preference while high doses of morphine (2.5, 5, 7.5 and 10 mg/kg) had a significant effect on the CPP (p<0.05, p<0.001, p<0.01 and p<0.001 respectively). Based on these studies 0.5 mg/kg (ineffective dose) and 5 mg/kg (effective dose) of morphine were selected for the subsequent experiments (Figure 2). Also, our results showed that morphine reward conditioning in low doses on the contrary to the high doses was dose dependent (Figure 2).
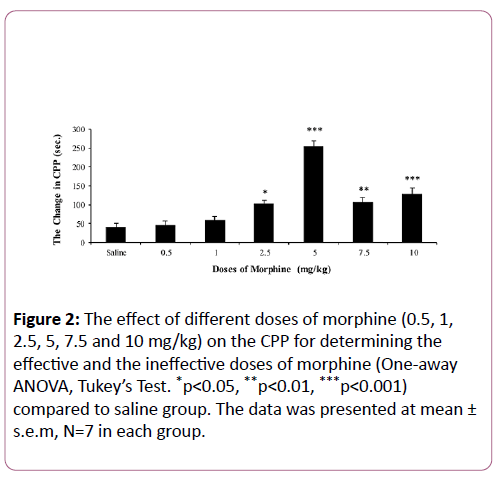
Figure 2: The effect of different doses of morphine (0.5, 1, 2.5, 5, 7.5 and 10 mg/kg) on the CPP for determining the effective and the ineffective doses of morphine (One-away ANOVA, Tukey’s Test. *p<0.05, **p<0.01, ***p<0.001) compared to saline group. The data was presented at mean ± s.e.m, N=7 in each group.
Microinjection of low-dose CNOX into the PL with different doses of morphine and its effect on the acquisition and the expression phases of the CPP paradigm
Analysis of our findings demonstrated that the microinjection of low dose of the CNQX (0.5 μg/0.5 μl) had significant effect on the acquisition phase of the CPP induced by the effective dose of morphine (p<0.001) but, this dose of the CNQX with the ineffective dose of morphine produced neither preference nor aversion effect on the acquisition phase of the CPP paradigm (p=0.063), Figure 3a.
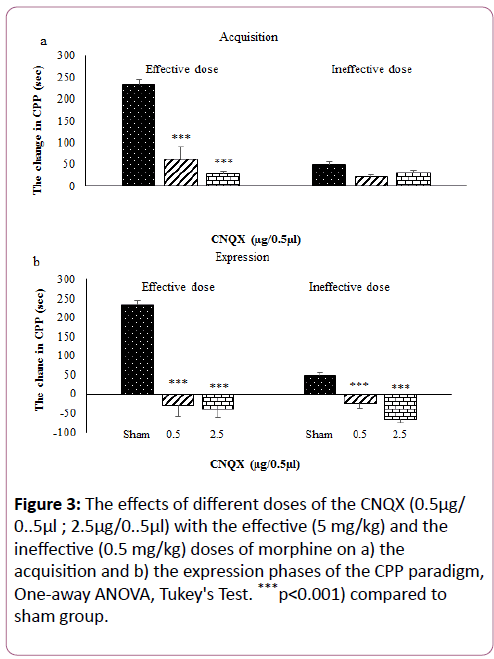
Figure 3: The effects of different doses of the CNQX (0.5μg/ 0..5μl ; 2.5μg/0..5μl) with the effective (5 mg/kg) and the ineffective (0.5 mg/kg) doses of morphine on a) the acquisition and b) the expression phases of the CPP paradigm, One-away ANOVA, Tukey's Test. ***p<0.001) compared to sham group.
Data analysis with a post-hoc test showed that the microinjection of low-dose CNQX into the PL with the effective dose of morphine, significantly reduced the expression phase of the CPP paradigm compared to the sham group (p<0.001). Also, this dose of the CNQX with the ineffective dose of morphine significantly induced an aversion effect on the expression phase of the CPP paradigm (p<0.001), Figure 3b.
Microinjection of high-dose CNQX into the PL with different doses of morphine and its effect on the acquisition and the expression phases of the CPP paradigm
The statistical analysis of ANOVA showed that the microinjection of high-dose CNQX (2.5 μg/0.5 μl) into the PL significantly decreased the acquisition phase of the CPP induced by the effective dose of morphine compared to the sham group (p<0.001). The microinjection of high-dose CNQX into the PL with ineffective dose of morphine produced neither preference nor aversion effect on the acquisition phase of the CPP (p=0.541), Figure 3a.
Our results showed that the microinjection of high-dose CNQX into the PL with the effective or the ineffective doses of morphine significantly induced not only reverse but also aversion effect on the expression phase of the CPP compared to sham group (p<0.001), Figure 3b.
The effect of the CNQX on drug-naïve and morphine-conditioned animals
Our findings indicated intra-PL different doses of the CNQX versus intra-PL saline produced no effect on drug-naïve animals (p=0.100), Figure 4a. Statistical analysis on difference score from baseline to test sessions can be performed to determine effects of drug conditioning. Therefore, based on statistical analysis difference score between intra-PL saline and intra-PL CNQX groups showed a significant effect on times spent in morphine-paired environment relative to saline-paired environment (p<0.001), Figure 4b. These results showed significant effect of CNQX on morphine-conditioned while had no effect on drug-naïve animals.
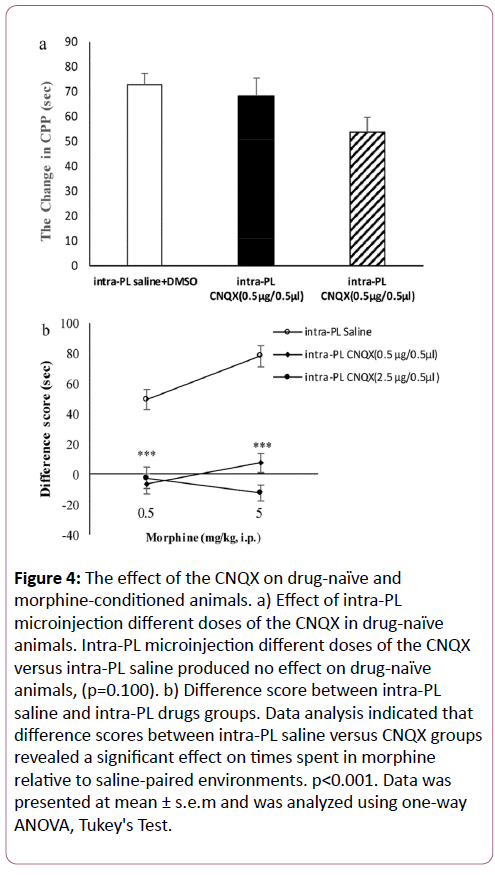
Figure 4: The effect of the CNQX on drug-naïve and morphine-conditioned animals. a) Effect of intra-PL microinjection different doses of the CNQX in drug-naïve animals. Intra-PL microinjection different doses of the CNQX versus intra-PL saline produced no effect on drug-naïve animals, (p=0.100). b) Difference score between intra-PL saline and intra-PL drugs groups. Data analysis indicated that difference scores between intra-PL saline versus CNQX groups revealed a significant effect on times spent in morphine relative to saline-paired environments. p<0.001. Data was presented at mean ± s.e.m and was analyzed using one-way ANOVA, Tukey's Test.
Locomotor activity
Our results showed that, intra-PL microinjection of difference doses of the CNQX with the effective dose and the ineffective dose of morphine had not significant effect on locomotor activity in the CPP paradigm in morphine-free state (P=0.7768, p=0.066 respectively), Figure 5.
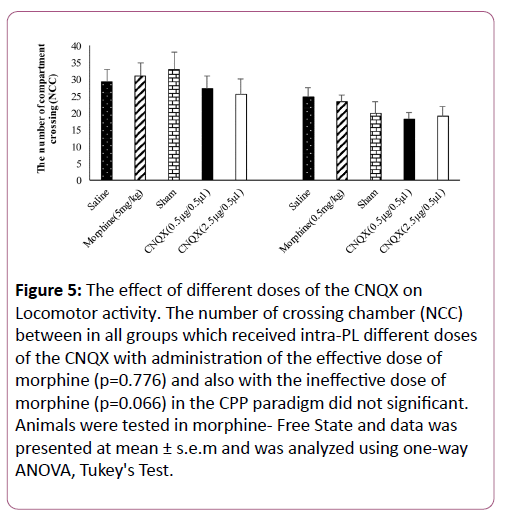
Figure 5: The effect of different doses of the CNQX on Locomotor activity. The number of crossing chamber (NCC) between in all groups which received intra-PL different doses of the CNQX with administration of the effective dose of morphine (p=0.776) and also with the ineffective dose of morphine (p=0.066) in the CPP paradigm did not significant. Animals were tested in morphine- Free State and data was presented at mean ± s.e.m and was analyzed using one-way ANOVA, Tukey's Test.
Discussion
In this study, different doses of morphine were tested according to the CPP for determining the effective and the ineffective dose of it. Our findings showed that the high doses of morphine (2.5, 5, 7.5 and 10 mg/kg) induced place preference, but low doses (0.5 and 1 mg/kg) had no effect on the CPP (Figure 2). In agreement with our findings, Kargari et al. and Ghavipanjeh GR et al. reported that systemic administration of morphine induced CPP and Van Der Kooy et al. also showed that unilateral injection of morphine into the NAc and VTA produced CPP in rats [1].
According to other study, it is possible that morphine induced these effects via the subunits of the AMPA receptors at glutamatergic synapses in dopamine neurons and changes activity of the mesolimbic dopamine system [26].
In other set of our experiments, results showed that intra-PL microinjection of the CNQX before administration of the effective dose of morphine significantly decreased the acquisition and the expression phases of the CPP (Figure 3). Similar to these results, injection of the CNQX inhibited development of cocaine sensitization. Data suggested that behavioral sensitization is associated with glutamate transmission via the AMPA receptors in the NAc which allows for increasing glutamate release in this region [27]. Other people reported that modulation activity of the AMPA receptor is critically involved in synaptic plasticity which is facilitated learning related-morphine [4]. On the other hand, the blockade of glutamate AMPA receptor with the CNQX inhibits produce preference in the CPP [28]. One possibility mechanism is that microinjection of glutamate antagonists into the PL may alter morphine-related dopaminergic signals from the VTA to the nucleus accumbens (NAc) and or change activity associative reward signals that converging from mPFC and VTA on the NAc [13]. Accordingly, the blockade of glutamate receptors in the PL disrupts the pathway between the prefrontal cortex, VTA and NAc that will alter dopaminergic neural activity and function of the reward system [29].
Our results also, revealed that the microinjection of different doses of the CNQX into the PL before the administration of the ineffective doses of morphine produced neither preference nor aversive effect in the acquisition phase of the CPP (Figure 3). Probably, glutamate balance plays an important role in opiate-conditioned memories. Data suggested that interaction between antagonist and balance in glutamate homeostasis [30] may alter the level of glutamate available for binding its receptors that prevents from change in the acquisition phase of the CPP with the ineffective dose of morphine [9]. Therefore, it seems that the interaction between the glutamate level and the antagonist did not attenuate the development of the morphine-ineffective dose in the acquisition phase of the CPP (Figure 3).
Recent study also found that the microinjection of different doses of the CNQX into the PL reversed the effect of the morphine-ineffective dose in the expression phase of the CPP (Figure 3b), which can be due the fact that many other brain regions may respond to reward systems [25,31] during conditioning of natural rewards with the exposure to addictive drugs. Probably, the effect of glutamate antagonists on the expression phase of the CPP with the ineffective dose of morphine can be due to synergism effects of these agents or the activation of another neural pathways that can change the CPP [17].
Conclusion
The present study showed that microinjection different doses of the CNQX into the PL with effective dose of morphine decreased acquisition and expression phases of the CPP. One possibility mechanism is that microinjection of the CNQX into the PL may alter morphine-related dopaminergic signals from the VTA to the NAc. Therefore, the blockade of glutamate AMPA receptors with CNQX in the PL disrupts the pathway between the prefrontal cortex, VTA and NAc that will change dopaminergic neural activity and function of the reward system and memory process. In contrast, different doses of the CNQX with the ineffective dose of morphine only reduced the expression phase of the CPP, suggesting that this response due to the modulation of reward system by another neuronal circuits in the brain. Therefore, it seems that AMPA receptor transmission into the PL modulates opiate-related reward signalling and memory processing.
Acknowledgment
We would like to thank Dr. Ghavipanjeh and Dr. Radahmadi for their assistance during this protocol.
References
- Zarrindast MR, Rezayof A (2010) Morphine-Induced Place Preference: Interactions with Neurotransmitter Systems. Iran J Pharmaceutical Res 6: 3-15.
- Zakharov D, Lapish C, Gutkin B, Kuznetsov A (2016) Synergy of AMPA and NMDA receptor currents in dopaminergic neurons: a modeling study. Front ComputNeurosci 10: 1-11.
- Corcoran KA, Quirk GJ (2007) Activity in prelimbic cortex is necessary for the expression of learned, but not innate, fears. J Neurosci 27: 840-844.
- Gass JT, Chandler LJ (2013) The Plasticity of extinction: contribution of the prefrontal cortex in treating addiction through inhibitory learning. Front Psychiatry 4: 1-12.
- Morgane PJ, Galler JR, Mokler DJ (2005) A review of systems and networks of the limbic forebrain/limbic midbrain. Progneurobiol 75: 143-60.
- Volkow N, Baler R (2013) Addiction: A Disease of Self-Control. Neurosciences and the Human Person: New Perspectives on Human Activities Pontifical Academy of Sciences 1-7.
- Beneyto M, Meador-woodruff JH (2003) AMPA and NMDA associated postsynaptic protein expresion in the human dorsolateral prefrontal cortex. Ann NY AcadSci 1003: 352-355.
- Olive MF (2009) Metabotropic glutamate receptor ligands as potential therapeutics for addiction. CurrDrug Abuse Rev 2: 83-94.
- Peters J, De Vries TJ (2012) Glutamate mechanisms underlying opiate memories. Cold Spring HarbPerspect Med 2: 1-10.
- Alaei H, Huotari M, Piepponen PT, Ahtee L, Hanninen O, et al. (2003) Morphine releases glutamate through AMPA receptor in the ventral tegmental area: A microdialysis study in conscious rats. Med J Islamic Repub Iran 17: 225-231.
- Todtenkopf MS, Parsegian A, Naydenov A, Neve RL, Konradi C, et al. (2006) Brain reward regulated by AMPA receptor subunits in nucleus accumbens shell. J Neurosci 26: 11665-11669.
- De Jaeger X, Bishop SF, Ahmad T, Lyons D, Ng GA, et al. (2013) The effects of AMPA receptor blockade in the prelimbic cortex on systemic and ventral tegmental area opiate reward sensitivity. Psychopharmacology (Berl) 225: 687-695.
- Bishop SF, Lauzon NM, Bechard M, Gholizadeh S, Laviolette SR (2010) NMDA receptor hypofunction in the prelimbic cortex increases sensitivity to the rewarding properties of opiates via dopaminergic and amygd alar substrates. Cerebral Cortex 21:68-80.
- Wang B, Luo F, Zhang WT, HanCA JS (2000) Stress or drug priming induces reinstatement of extinguished conditioned place preference. NeuroReport 11:2781-2784.
- Ghalandari-Shamami M, Hassanpour-Ezatti M, Haghparast A (2012) Glutamate Receptors in Nucleus Accumbens Can Modulate Canabinoid-Induced Antinociception in Rat’s Basolateral Amygdala. Basic CliniNeurosci 3:11-18.
- Paxinos G, Watson C (2006) The Rat Brain in Stereotaxic Coordinates: Hard Cover Edition: Academic Press 456.
- CarlezonWA Jr, TodtenkopfMS, McPhie D, Pimentel P, Pliakas A, et al. (2001) Repeated exposure to rewarding brain stimulation downregulates GluR1 expression in the ventral tegmental area. Neuropsychopharmacology 25:234-241.
- Kargari A, Ramshini E, Alaei H, Sedighi M, Oryan S (2012) Different current intensities electrical stimulation of prelimbic cortex of mPFC produces different effects on morphine-induced conditioned place preference in rats. Behav brain res 231:187-192.
- Rahimpour M, Karami M, Karimi S, Haghparast A, Jalali MR, et al. (2011) Blockade of the Naloxone-induced Aversion in Morphine-conditioned Wistar Rats by L-Arginine Intra-central Amygdala. Iran J Basic Med Sci 14:167-176.
- Zarrindast MR, Asadi F, Rezayof A (2011) Repeated pretreatment of morphine prevents morphine-induced amnesia: A possible involvement for dorsal hippocampal NMDA receptors. Arch Iran Med 14:32-38.
- Zarrindast MR, Lashgari R, Rezayof A, Motamedi F, Nazari-Serenjeh F (2007) NMDA receptors of dorsal hippocampus are involved in the acquisition, but not in the expression of morphine-induced place preference. Eur JPharmacol 568:192-198.
- Ghavipanjeh GR, Pourshanazari AA, Alaei H, Karimi S, Nejad MA (2015) Effects of Temporary Inactivation and Electrical Stimulation of the Dorsal Raphe Nucleus on Morphine-induced Conditioned Place Preference. Malays JMed Sci 22:33-40.
- Bardo MT, Bevins RA (2000) Conditioned place preference: what does it add to our preclinical understanding of drug reward? Psychopharmacology (Berl) 153:31-43.
- Feduccia AA, Duvauchelle CL (2010) Novel Apparatus and Method for Drug Reinforcement. J Vis Exp 1-3.
- Dong Z, Han H, Wang M, Xu L, Hao W, et al. (2006) Morphine Conditioned Place Preference Depends on Glucocorticoid Receptors in Both Hippocampus and Nucleus Accumbens. Hippocampus 16:809-813.
- Brown MT, Bellone C, Mameli M, Labouèbe G, Bocklisch C, et al. (2010) Drug-driven AMPA receptor redistribution mimicked by selective dopamine neuron stimulation. PloS one 5:1-10.
- Steketee JD, Kalivas PW (2011) Drug wanting: behavioral sensitization and relapse to drug-seeking behavior. Pharmacological rev 63:348-365.
- Watanabe T, Nakagawa T, Yamamoto R, Maeda A, Minami M, et al. (2002) Involvement of Glutamate Receptors Within the Central Nucleus of the Amygdala in Naloxone-Precipitated Morphine Withdrawal-Induced Conditioned Place Aversion in Rats. Jpn J Pharmacol 88:399-406.
- Stuber GD, Hopf FW, Tye KM, Chen BT, Bonci A (2010) Neuroplastic alterations in the limbic system following cocaine or alcohol exposure.Behavioral Neuroscience of Drug Addiction: Springer 3: 3-27.
- Kalivas PW (2009) The glutamate homeostasis hypothesis of addiction. Nat Rev Neurosci 10:561-572.
- Ebrahimi-ghiri M (2012) The Effect of Cholestasis on Rewarding and Exploratory Behav-iors Induced by Opioidergic and Dopaminergic Agents in Mice. Arch Iran Med 15:617-624.