Cheng Peng* and Jack C Ng
National Research Centre for Environmental Toxicology (Entox), Queensland Alliance for Environmental Health Science (QAEHS), The University of Queensland, Coopers Plains, Brisbane, QLD 4108, Australia
*Corresponding Author:
Cheng Peng
Faculty of Health and Behavioural Sciences
Entox (National Research Centre for Environmental Toxicology) The University of Queensland
39 Kessels Road Coopers Plains, Brisbane, Queensland, Australia.
Tel: +61 429 547 155
Fax: +61 732739003
E-mail: c.peng@uq.edu.au
Received date: April 28, 2016; Accepted date: May 10, 2016; Published date: May 17, 2016
Citation: Peng C, Ng JC. The Role of Epigenetic Changes in Benzene-Induced Acute Myeloid Leukaemia. J Clin Epigenet. 2016, 2:2. DOI: 10.21767/2472-1158.100019
Copyright: © 2016 Peng C, Ng JC. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Acute myeloid leukaemia (AML) is a malignant disease of the blood and bone marrow system. Normally, different types of mature blood cells are derived from their progenitor cells and haematopoietic stem cells (HSC). In the haematopoietic system, the fate of the cells such as quiescence, self-renewal or proliferation, and differentiation at the progenitor or precursor level are strictly regulated. This haematopoietic hierarchy is controlled via a variety of growth factors including colony-stimulating factors (CSFs), interleukins (ILs), transcriptional factors, hormones and other regulation factors such as p53 and apoptotic process [1]. It has been established that uncontrolled proliferation, blocked differentiation and dysregulated apoptosis of haematopoietic precursor cells are involved in AML development, but the origin of the etiological changes in AML has not yet been completely elucidated. Some environmental carcinogens including benzene (Bz) can induce AML. Understanding the molecular mechanisms underlying Bz-induced leukaemogenesis will help to elucidate the initiation and progression of AML.
Bz Exposure and AML
It has been suggested that multiple key molecular and cellular events may be involved in myeloid leukaemogenesis. Exposure to environmental factors including chemicals (i.e., the leukaemogenic chemicals and some therapy-related compounds) as well as ionizing radiation is risk factors causing AML and myelodysplastic syndrome (MDS) [2]. Benzene (Bz), a volatile compound, is used in petrol and other manufactured products including rubber, lubricants, dyes, detergents and drugs. Additionally, Bz is an important air contaminant generated from motor vehicles, industrial processes, and cigarette smoking. Therefore, humans may be exposed to Bz through inhalation from a variety of different sources. Notably, Bz has been classified as a human carcinogen by the International Agency for Research on Cancer [3] and exposure to Bz and its association with AML has been well reviewed and documented [2]. After exposure, Bz undergoes metabolism by cytochrome P450 2E1 and 2F1 (CYP2E1 and CYP2F1) mainly in the liver. It has been well established that Bz exerts its toxicity through the reactive metabolites, i.e., catechol, hydroquinone (HQ), benzoquinone (BQ) and others [4]. Bz and its metabolites can redistribute and accumulate in bone marrow tissue where they exert their selective toxicity to the haematopoietic stem cells or progenitor cells. The resultant molecular and cellular effects include oxidative stress, changes in gene expression and function, disrupted balance among proliferation, differentiation, apoptosis and cytogenetic abnormalities like chromosomal aberrations [5]. Recently, epigenetic changes induced by Bz and their roles in disease initiation and development have drawn great attentions.
Environmental Factor-Induced Epigenetic Changes
Epigenetic refers to inheritable changes in phenotype and function without alteration of DNA sequences [6]. In other words, epigenetic modulation regulates or modulates in some degree gene expression and consequent functions. This means that normal physiological functions of cells are controlled by not only genetic mechanisms but also balanced epigenetic pattern which plays an important role in the control of many cellular functions of the body. In addition, environmental factors can cause epigenetic modifications through various mechanisms. In addition, many genetic changes induced by environmental factors such as environmental chemicals are virtually through epigenetic modifications. One of the mechanisms of the environmental chemical-induced epigenetic alterations is due to the generation of reactive oxygen species (ROS) [7, 8]. The environmental factor-induced epigenetic changes and consequent health effects have drawn great attentions in recent years in clinical and health research areas. The epigenetic modifications include DNA methylation, histone modifications and non-coding RNAs (ncRNA) expression. Methylation of DNA, a process involving the addition of methyl groups to DNA typically at CpG dinucleotide context, can cause the conformational change of DNA structure and consequent alteration in gene expression [9, 10]. DNA methylation is import regulation mechanism for mammalian development [11]. However, abnormal DNA methylation patterns, hypermethylation or hypomethylation can lead to various pathogenesis or oncogenesis. Hypermethylations are generally associated with gene silencing or down regulation, whereas hypomethylation or unmethylated promoters are mostly linked to gene activation [12]. Epigenetic regulation of gene expression can also be through modification of histone through posttranslational modifications such as acetylation, phosphorylation, methylation and ubiquitination [13]. Histone modifications are important in genetic process including transcriptional regulation, DNA repair, DNA replication, alternative splicing and chromosome condensation [13-15]. Another important epigenetic modifier is ncRNA including microRNAs (miRs) and long non-coding RNAs (LncRNAs) [16]. Many environmental chemicals including benzene can modify normal epigenetic pattern.
Bz-Induced Epigenetics Alterations and Association with AML
Although Bz has been well studied for its genotoxic and carcinogenetic effects in animal and human studies, the exact mechanisms underlying are still remained to be understood. Bz has been known to have genotoxic effects, but emerging evidence has suggested that this compound can also induce epigenetic changes. The reason of Bz-induced epigenetic modifications is unclear but ROS may be one of the pathways involved since Bz and its metabolites have been shown to generate ROS during metabolic transformation [17-19] and ROS can cause epigenetic changes as mentioned above. Further work is required to investigate the cause of Bz-induced epigenetic alterations, which will provide more mechanistic clues.
The epigenetic role in Bz-induced carcinogenesis has emerged only in recent years with some studies published. Although the epigenetic effect of Bz in progression, development and survival of the leukemic clone was proposed as early as in 1996 [20], recent studies suggested an earlier and broader involvement of epigenetic modification in Bz-induced hematotoxicity. Bollati V and his colleagues conducted the earlier human study of Bz exposure and associated epigenetic changes [21]. They found that exposure to Bz at low level can cause to significant reduction in long interspersed nuclear element-1 (line 1) and AluI repetitive elements methylation and hypomethylation in MAGE-1 and p15. The epigenetic alterations in these genes were also found in AML cells [22-25]. Hypomethylation of Melanoma-associated antigen 1 (MAGE-1) was further confirmed together with finding of another gene associated with AML, runt-related transcription factor 1 (RUNX1T1) in an in vitro study in which TK6 cells were treated with hydroquinone, one of Bz’s metabolites [26]. In addition, Bz induce methylation of PTEN (Phosphatase and tensin homolog deleted on chromosome 10), a tumor suppresser gene, leading to altered expression and function of this gene [27].
Furthermore, Bz was found to cause aberrant promoter methylation of genes for DNA repair such as ERCC1 [28] and PARP- 1 [29]. It has been believed Bz elicits its carcinogenesis through its metabolites which can directly or indirectly induce ROS leading to DNA damage. In this context, DNA repair plays important role in Bz-induced carcinogenesis and reduced DNA repair capacity may lead to increased susceptibility to Bz toxicity.
Bz may also elicit its hematotoxicity through increase the histone deacetylases (HDAC) [30]. Bz metabolites have been found to Methyl-CpG-binding domain protein 2 (MBD2) and DNA methyltransferases (DNMTs) were found to be hypomethylated by benzene exposure [31]. Among the epigenetic modifiers, HDAC and DNMTs (DNMT3A) have been showed to associate with the early stage of AML [32-34]. Therefore, the dysfunction of these epigenetic modifiers by Bz and its metabolites may further contribute to its carcinogenesis at initiation and development stage.
In conclusion, recent epigenetic studies on Bz although not many suggested that Bz can induce a variety of epigenetic alterations on genes involving in tumor suppresser (p15 and PTEN), cancer antigen (MAGE-1), transcription (RUNX1T1), DNA repair (PARP- 1, ERCC-1), and even epigenetic modification (HDAC and DNMTs) as summarized in Figure 1. Importantly, some of these genes are found to be modified epigenetically in AML cells which may indicate the role of these epigenetics alterations in Bz-induced leukaemogensis of which further research work will provide more mechanistic information.
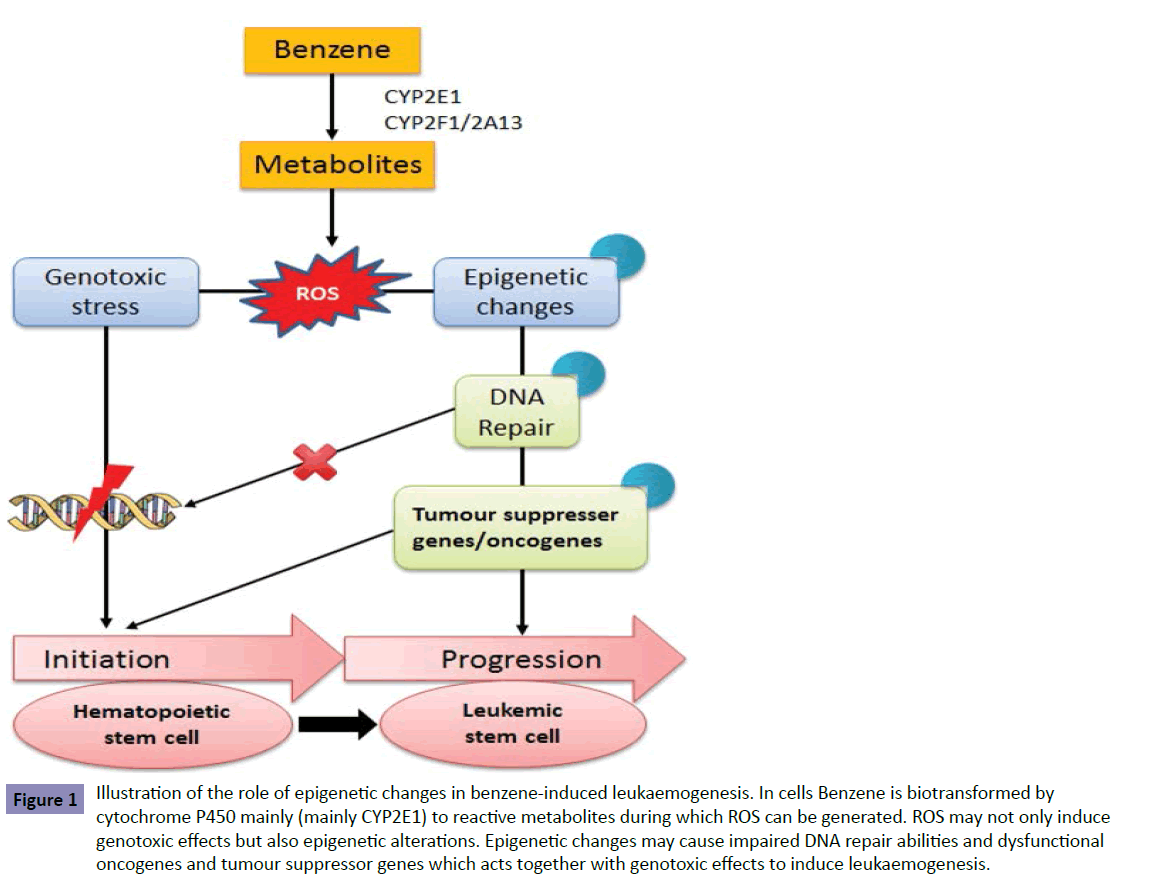
Figure 1: Illustration of the role of epigenetic changes in benzene-induced leukaemogenesis. In cells Benzene is biotransformed by cytochrome P450 mainly (mainly CYP2E1) to reactive metabolites during which ROS can be generated. ROS may not only induce genotoxic effects but also epigenetic alterations. Epigenetic changes may cause impaired DNA repair abilities and dysfunctional oncogenes and tumour suppressor genes which acts together with genotoxic effects to induce leukaemogenesis.
References
- Sive JI, Gottgens B (2014) Transcriptional network control of normal and leukaemic haematopoiesis. Exp Cell Res 329: 255-264.
- McHale CM, Zhang L, Smith MT (2012) Current understanding of the mechanism of benzene-induced leukemia in humans: implications for risk assessment. Carcinogenesis 33: 240-252.
- IARC (2012) Agents Classified by the IARC Monographs, IARC Monographs on the Evaluatio of Carcinogenic Risks to Human. 29: 100F.
- Ross D (2000) The role of metabolism and specific metabolites in benzene-induced toxicity: Evidence and issues. Journal of Toxicology and Environmental Health 61: 357-372.
- Greim H, Kaden DA, Larson RA, Palermo CM, Rice JM, et al. (2014) The bone marrow niche, stem cells, and leukemia: impact of drugs, chemicals, and the environment. Ann N Y AcadSci 1310: 7-31.
- Gibney ER, Nolan CM (2010) Epigenetics and gene expression. Heredity 105: 4-13.
- Ziech D, Franco R, Pappa A, Malamou-Mitsi V, Georgakila S, et al. (2010) The role of epigenetics in environmental and occupational carcinogenesis. ChemBiol Interact 188: 340-349.
- Ziech D, Franco R, Pappa A, Panayiotidis MI (2011) Reactive oxygen species (ROS)--induced genetic and epigenetic alterations in human carcinogenesis. Mutat Res 711: 167-173.
- Prokhortchouk E, Defossez PA (2008) The cell biology of DNA methylation in mammals. BiochimBiophysActa 1783: 2167-2173.
- Cao JX, Zhang HP, Du LX (2013) Influence of environmental factors on DNA methylation. Yi chuanHereditas 35: 839-846.
- Smith ZD, Meissner A (2013) DNA methylation: roles in mammalian development. Nat Rev Genet 14: 204-220.
- Laffita-Mesa JM, Bauer P (2014) Herenciaepigenética (metilacióndelácidodesoxirribonucleico): contextoclínico en neurodegeneraciones y gen ATXN2. MedicinaClínica 143: 360-365.
- Kouzarides T (2007) Chromatin modifications and their function. Cell 128: 693-705.
- Huertas D, Sendra R, Munoz P (2009) Chromatin dynamics coupled to DNA repair. Epigenetics 4: 31-42.
- Luco RF, Pan Q, Tominaga K, Blencowe BJ, Pereira-Smith OM et al. (2010) Regulation of alternative splicing by histone modifications. Science 327: 996-1000.
- Peschansky VJ, Wahlestedt C (2014) Non-coding RNAs as direct and indirect modulators of epigenetic regulation. Epigenetics 9: 3-12.
- Gut I, Nedelcheva V, Soucek P, Stopka P, Tichavska B (1996) Cytochromes P450 in benzene metabolism and involvement of their metabolites and reactive oxygen species in toxicity. Environmental Health Perspectives 104: 1211-1218.
- Shen Y, Shen HM, Shi GY, Ong GN (1996) Benzene metabolites enhance reactive oxygen species generation in HL60 human leukemia cells. Human and Experimental Toxicology 15: 422-427.
- Peng C, Arthur D, Liu FY, Lee J, Xia Q, et al. (2013) Genotoxicity of hydroquinone in A549 cells. Cell Biology and Toxicology 29: 213-227.
- Irons RD, Stillman WS (1996) The process of leukemogenesis. Environ Health Perspect 104: 1239-1246.
- Bollati V, Baccarelli A, Hou A, Bonzini M, Fustinoni M, et al. (2007) Changes in DNA methylation patterns in subjects exposed to low-dose benzene. Cancer Res 67: 876-880.
- Lubbert M, Oster W, Ludwig WD, Ganser A, Mertelsmann R, et al. (1992) A Switch toward Demethylation Is Associated with the Expression of Myeloperoxidase in Acute Myeloblastic and PromyelocyticLeukemias. Blood 80: 2066-2073.
- Melki JR, Vincent PC, Clark SJ (1999) Concurrent DNA hypermethylation of multiple genes in acute myeloid leukemia. Cancer Res 59: 3730-3740.
- Greiner J, Ringhoffer M, Simikopinko O, Szmaragowska A, Huebsch S, et al. (2000). Simultaneous expression of different immunogenic antigens in acute myeloid leukemia. Experimental Hematology 28: 1413-1422.
- Galm O, Herman JG, Baylin SB (2006) The fundamental role of epigenetics in hematopoietic malignancies. Blood Rev 20: 1-13.
- Zhang LP, McHale CM, Rothman N, Li GL, Ji ZY, et al. (2010) Systems biology of human benzene exposure. Chemico-Biological Interactions 184: 86-93.
- Yang J, Zuo X, Bai W, Niu P, Tian L, et al. (2014) PTEN methylation involved in benzene-induced hematotoxicity. ExpMolPathol 96: 300-306.
- Xing CH, Chen Q, Li GL, Zhang LY, Zheng M, et al. (2013). Microsomal epoxide hydrolase (EPHX1) polymorphisms are associated with aberrant promoter methylation of ERCC3 and hematotoxicity in benzene-exposed workers. Environmental and Molecular Mutagenesis 54: 397-405.
- Gao A, Zuo X, Liu JQ, Lu X, Guo W, et al. (2010). Methylation of PARP-1 promoter involved in the regulation of benzene-induced decrease of PARP-1 mRNA expression. Toxicology Letters 195: 114-118.
- Chen J, Zheng Z, Chen Y, Li J, Qian S, et al. (2016) Histone Deacetylase Inhibitors Trichostatin A and MCP30 Relieve Benzene-Induced Hematotoxicity via Restoring Topoisomerase IIalpha. PLoS One 11: e0153330.
- Sha Y, ZhouW, YangZ, ZhuX, XiangY, et al. (2014) Changes in poly(ADP-ribosyl)ation patterns in workers exposed to BTX. PLoS One 9: e106146.
- Mizuno S, Chijiwa T, Okamura T, Akashi K, Fukumaki Y, et al. (2001) Expression of DNA methyltransferases DNMT1, 3A, and 3B in normal hematopoiesis and in acute and chronic myelogenousleukemia. Blood 97: 1172-1179.
- Tickenbrock L, Klein HU, Trento C, Hascher A (2011) Increased HDAC1 deposition at hematopoietic promoters in AML and its association with patient survival. Leukemia Research 35: 620-625.
- Abdel-Wahab O, Levine RL (2013) Mutations in epigenetic modifiers in the pathogenesis and therapy of acute myeloid leukemia. Blood 121: 3563-3572.

